Inviato 06 giugno 2006 - 17:20
Osservazioni
Lappartenenza del genere Inocybe al gruppo trofico dei cosiddetti simbionti ectomicorrizici è universalmente accettata; sembra, peraltro, che la regola possa ammettere delle eccezioni, sussistendo la possibilità per certe specie di svolgere, in particolari circostanze, unesistenza saprofitica. La descrizione ecologica di I. rimosobispora, che viene data, almeno fino ad ora, nelle radure di querce da sughero, su residui legnosi decomposti più o meno interrati, potrebbe indurre la sensazione che essa possa essere definita come specie saprotrofa. Tuttavia ci limitiamo a descrivere le nostre osservazioni senza, per tale motivo, giungere a conclusioni avventate e, soprattutto, non dimostrabili.
Le cinque raccolte studiate, provenienti da tre diverse località della Sardegna, ci hanno mostrato una rimarchevole variabilità di questa specie nei caratteri macromorfologici: in particolare, il gambo può presentare o meno, alla sommità, una tenue sfumatura rosa essa era presente nelle raccolte di Stazzo Montesu ed assente, viceversa, nelle altre e le lamelle hanno uninserzione piuttosto variabile, potendo essere da annesse a subdecorrenti mentre la carne può presentare o meno un tenue odore spermatico.
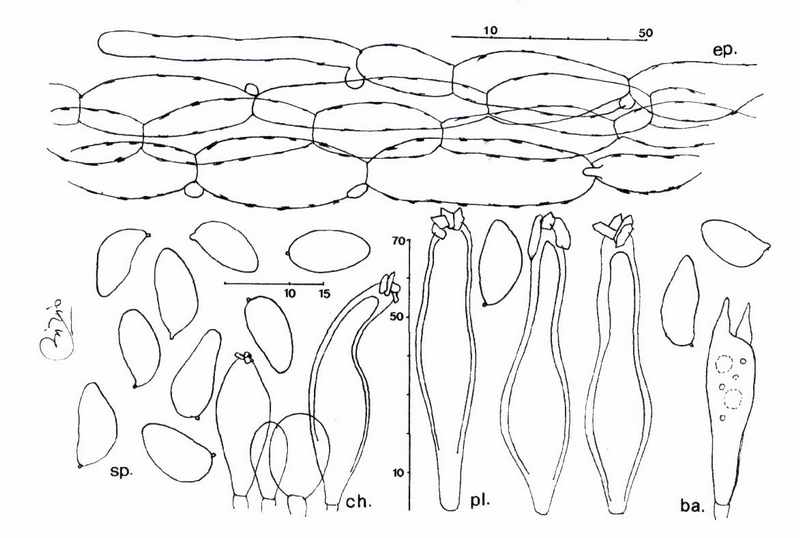
Dal punto di vista micromorfologico, tuttavia, I. rimosobispora appare piuttosto stabile, soprattutto se si ha riguardo alla bisporia nonché alla forma ed alle dimensioni di spore e cistidi.
Per le spore lisce ed i cistidi a parete spessa questa specie appartiene, secondo limpostazione sistematica di BON (1997), al sottogenere Inocybe e, in seno a questo, per lassenza di cortina, per le colorazioni banali, per lassenza di viraggi e per il gambo in gran parte pruinoso, non bulboso, alla sezione Splendentes R. Heim ex Singer. La successiva attribuzione ad una delle sottosezioni della sezione Splendentes risulta molto problematica per limbarazzo cui ci hanno posto alcuni basidiomi che presentavano una lieve sfumatura rosa sulla corteccia del gambo, più specificatamente nella loro parte superiore.
Con riguardo alle colorazioni del gambo, Bon definisce la sottosezione Phaeoleucinae Bon con il gambo non rosato e la sottosezione Subbrunneinae Bon con gambo +/- rosato. In questa seconda sottosezione, egli colloca parecchie specie con gambo inizialmente bianco, poi debolmente rosato. Tra queste spicca I. hirtella Bres. che, secondo il punto di vista del micologo francese, deve includere la var. bispora Kuyper.
Se usiamo dunque lo stesso criterio di valutazione di Bon, dovremmo collocare senzaltro I. rimosobispora tra le Subbrunneinae, però è chiaro che si tratta di una specie dotata di un grande dinamismo cromatico, tale da consentire lattribuzione ora alluno, ora allaltro dei due gruppi considerati. La difficoltà di definire il colore finisce per aumentare il rischio di darne una valutazione soggettiva, con i nefasti risultati che chiunque può immaginare. A tale proposito sorge spontanea unosservazione mossa dallesperienza: cercare di pianificare una suddivisione infragenerica (in Inocybe), basata sui colori del gambo, quando essi non siano decisi, è unoperazione illusoria; la realtà, in effetti, non è sempre così rigida. La strada intrapresa da Kühner (KÜHNER & ROMAGNESI, 1953) che cercò di separare i gruppi con tinte rosa da quelli con tinte gialle ha destato un certo fascino nei micologi successivi, tanto che essa è tuttoggi esplorata da non pochi studiosi del genere. Tuttavia, una parallela scuola di pensiero, ritiene tale separazione cromatica ingiustificata se non addirittura artificiale, appartenendo (probabilmente) questi due raggruppamenti ad un medesimo gruppo naturale (KUYPER, 1986). E emblematico che tra le specie affini di I. rimosobispora vi siano entità (le già citate I. amygdalispora e I. hirtella var. bispora) altalenanti (a seconda della soggettività) tra le sottosezioni Phaeoleucinae e Subbrunneinae, che sono raggruppamenti, come già detto, separati da una presunta differenza cromatica.
Microscopicamente, I. rimosobispora deve essere posta in stretta correlazione con I. descissa var. bisporigera J.E. Lange. Nella tav. 200, figura F, il micologo danese (LANGE, 1940) rappresenta unInocybe che viene descritta (invalidamente ex I.C.B.N., art. 36.1) con cappello rimoso verso lorlo, il gambo bruniccio pallido, minutamente pruinoso, specie in alto; essa è dotata di basidi bisporici. Si tratta di una entità molto controversa: KUYPER (1986) che ne ha analizzato il typus, lha ricondotta ad una varierà bisporica di I. fuscidula Vel. (specie anchessa assai problematica).Ma listituzione di I. fuscidula var. bisporigera da parte di Kuyper è una operazione che ha lasciato non poche perplessità: si è osservato che I. fuscidula è specie a gambo bianco o biancastro sporco, mai dotata della colorazione descritta da Lange. Per questo motivo essa è collocata nella sottosezione Virgulatinae Bon, gruppo nel quale confluiscono le specie a cauloimenio parziale e, per lappunto, senza tracce di colorazioni rosate sul gambo. Ad ogni buon conto, I. fuscidula var. bisporigera differisce molto bene da I. rimosobispora per la presenza di velo e cortina, per il gambo provvisto di caulocistidi solo nel terzo superiore e per i cistidi mediamente più larghi.
I. rimosobispora potrebbe anche essere vista come una manifestazione bisporica di I. pseudodestricta Stangl & J. Veselsky, alla quale molto assomiglia per laspetto macromorfologico. Il caso ha voluto che proprio uno di noi si sia imbattuto in passato in una manifestazione bisporica di questa specie (Maniago, 28.05.1994, legit E. B., sotto Fraxinus excelsior). Questultima però differisce da I. rimosobispora per un certo numero di elementi: il cappello uniformemente rosso-bruno, la cuticola pileica più brillante, fibrillosa, poco o affatto rimosa, la sfumatura giallastra delle lamelle, il gambo più robusto, bianco-ocraceo e intensamente rosa-lilla allapice, con base bianca, i cistidi imeniali più corti e larghi, i caulocistidi presenti limitamente al primo sesto superiore. Sfortunatamente, finora, possiamo disporre di una sola raccolta di questa variante bisporica di I. pseudodestricta; questa circostanza ci impone cautela e ci induce ad attendere eventuali nuovi ritrovamenti per istituire ufficialmente la nuova varietà. I. pseudodestricta, nella concezione di Bon, col nome di I. destricta (Fr.) Quél. (= I. pseudodestricta Stangl & J. Veselsky ss. Kuyper) è collocata nella sottosezione Tardinae Bon (specie con caulocistidi nel terzo superiore del gambo e colorazioni rosate in alcune parti del carpoforo) non lontana da I. nitidiuscula (Britzelm.) Sacc (=. I. friesii R. Heim).
Leventualità di imbattersi in basidiocarpi del genere Inocybe provvisti di basidi bisporici è, a tutti gli effetti, piuttosto infrequente. Vi sono numerose specie che possono presentare basidi con (1), 2 o 4 spore, tuttavia esse sono solitamente associate a particolari situazioni ecologico-fenologico estreme (BIZIO, 1997): ambienti xerico/sabulicoli, zona alpina, stagione invernale. Solo I. rimosobispora e le due varietà bisporiche di I. fuscidula Velen. e di I. hirtella Bres. possiedono basidi sostanzialmente o esclusivamente bisporici, almeno tra tutte quelle descritte per lItalia e per lEuropa.
BON (1999) ha descritto, ad interim, una varietà bisporica di I. ovalispora C.H. Kauffm. Anche questa raccolta, della quale non si hanno ulteriori notizie, presenta singolari analogie con I. rimosobispora: il cappello rosso-bruno, rimoso e la crescita su frammenti legnosi interrari di Quercus ilex. Tuttavia, vi sono numerosi elementi che separano nettamente le due entità: gli esemplari di Bon possiedono il gambo già inizialmente e completamente di colore rosa, un bulbo marginato alla base, lodore leggermente di pelargonio, le spore in maggioranza con apice ottuso, i cistidi con parete nettamente più sottile. I. ovalispora C.H. Kauffm. è una specie originaria del Michigan (U.S.A.) diversamente interpretata dagli autori europei, tanto da essere di volta in volta relazionata ad un gran numero di specie diverse, seppure affini: I. albomarginata Velen., I. tjallingiorum Kuyper, I. catalaunica Sing. (= I. subbrunnea Kühner), la f. tenerella Favre di I. ovalispora [più tardi I. tenerella (Favre) ex Kuyper & Month. ined.]; al binomio ovalispora-subbrunnea sono state successivamente ricondotte le forme brunneola e tenerella del Favre. Lo studio del typus di Kauffman da parte di KUYPER (1986) ha stabilito la conspecificità di I. ovalispora e I. albomarginata Velen., pubblicata nel 1920. La combinazione di caratteri: gambo rosa e bulbo +/- marginato ha permesso a BON (1997) listituzione di una stirpe Ovalispora inclusa nella sottosezione Splendentinae.
Ad una approssimativa osservazione, i basidi bisporici e lassenza di velo potrebbero favorire confusioni anche con la già citata I. hirtella var. bispora Kuyper (1986 ) ma questa entità differisce dalla nostra per il cappello scarsamente umbonato e di colorazione bruno-giallastra, la carne con tipico odore di mandorle amare ed i cistidi più corti e decisamente più larghi, di profilo più tozzo. Secondo Kuyper, sulla scorta dello studio derbario, la varietà bisporici di I. hirtella Bres. deve includere I. amygdalispora, specie a basidi bisporici invalidamente (art. 36.1 I.C.B.N.) descritta da Métrod nel 1956. Tuttavia, e ciò sia detto per inciso, è allo studio di uno di noi (M.C.) il confronto tra le due specie: tale studio si propone di valutare la possibilità di mantenere separate le due entità.
Infine, dopo le varietà bisporiche di I. fuscidula e di I. hirtella, possiede basidi bisporici, secondo la diagnosi originale, una terza specie europea: si tratta di I. ovoideicystis Métrod nom. inval. (METROD, 1956, BON, 1997), da Kuyper sinonimizzata ad I. appendiculata Kühner (KUYPER, 1986), entità a cistidi di foggia e dimensioni completamente diverse (ovoidali o piriformi, lunghi fino a 55 µm e larghi fino a 20 µm) e spore alquanto minori, di profilo ellissoidale.
Se allarghiamo lindagine alle specie extraeuropee, troviamo descritto un limitatissimo numero di specie, tassonomicamente molto distanti dalla specie qui proposta come nuova. Infatti:
I. aberrans (E. Horak) Garrido (HORAK, 1979 sub Astrosporina aberrans), specie delle Isole Papua Nuova Guinea, possiede spore nodulose; è da ascrivere alla sezione Cortinatae Kühner & Boursier.
I. bispora Hongo (1958), dal Giappone, pur caratterizzata da spore lisce, ha cistidi a parete sottile e per di più presenti solo sul taglio lamellare; è una tipica rappresentante della sezione Rimosae (Fr.) Sacc.

 Questa discussione è bloccata
Questa discussione è bloccata