RITROVAMENTI FUNGINI IN AMBIENTI ANTROPIZZATI DELLA VENEZIA GIULIA
II PARTE
di Paolo Picciola* e Marino Zugna**
(*) Paolo Picciola – Strada di Fiume, 52 - 34137 Trieste (TS) paolo.picciola2@gmail.com
(**) Marino Zugna - Località Rio Storto, 7 - 34015 Muggia (TS) marinozugna@gmail.com
Introduzione
In questo secondo contributo gli autori descrivono, commentano e documentano con immagini dei basidiomi in habitat e dei caratteri microscopici essenziali, ulteriori undici specie fungine rinvenute negli ambienti urbani della Venezia Giulia. Sono presentati sia taxa considerati comuni e tipici degli ambienti antropizzati, facilmente riconoscibili con l’osservazione dei soli caratteri macroscopici, sia taxa rari o poco comuni per il cui riconoscimento è necessario ricorrere a uno studio completo dei caratteri microscopici.
La maggior parte delle raccolte presentate sono state effettuate nella zona sud-est della città di Trieste, nei quartieri di Ponziana e Campi Elisi e nel comune di Muggia.
Materiali e metodi
La descrizione dei caratteri macroscopici è stata ricavata analizzando i basidiomi freschi fotografati in habitat mediante fotocamere digitali Nikon D90 (P.P.) e Canon EOS 5D Mark II (M.Z.).
Lo studio dei caratteri microscopici è stato eseguito sia sul fresco sia su materiale d’erbario, in quest’ultimo caso reidratato in acqua distillata o una soluzione acquosa d’idrossido di potassio 5%, e, ove necessario, colorato con rosso Congo ammoniacale 2% per meglio evidenziare le differenti tipologie cellulari.
Le foto concernenti la microscopia sono state eseguite con l’ausilio di una fotocamera Reflex Canon EOS 50D.
Le misurazioni microscopiche sono state eseguite con il software di calcolo Mycométre (http://mycolim.free.fr).
Per la terminologia riguardante la tipologia delle caratteristiche macro e microscopiche si fa riferimento a Vellinga (1988).
Per quanto concerne la nomenclatura delle specie presentate si è fatto riferimento alle indicazioni contenute nei siti http://www.indexfungorum.org/ e http://www.mycobank.org. Gli exsiccata sono depositati nell'erbario del gruppo micologico A.M.B. di Muggia e del Carso.
Specie fungine rappresentate
Vengono descritte e commentate le seguenti 11 specie fungine:
1. Pholiotina dasypus
2. Conocybe macrocephala
3. Coprinopsis marcescibilis
4. Parasola conopilus
5. Parasola auricoma
6. Panaeolus cinctulus
7. Psilocybe coronilla
8. Volvopluteus gloiocephalus
9. Callistosporium olivascens
10. Cyclocybe cylindracea
11. Lepiota subincarnata
Pholiotina dasypus
(Romagn.) P.-A. Moreau, Fungal Diversity 20: 135 (2005)
Basionimo: Naucoria dasypus Romagn., Bulletin de la Société Mycologique de France 53: 121 (1937)
Sinonimi: Alnicola dasypus (Romagn.) Romagn., Bulletin de la Société Mycologique de France 58
(4): 126 (1942). Hebeloma dasypus (Romagn.) Singer, Lilloa 22: 538 (1951). Conocybe subnuda Kühner, Encyclopédie Mycologique 7: 140 (1935). Conocybe utriformis P.D. Orton, Transactions of the British Mycological Society 43 (2):196 (1960). Conocybe subverrucispora Veselský & Watling, Ceská Mykologie 26 (4): 201 (1972).
Conocybe subnuda Kühner ex Kühner & Watling, Notes from the Royal Botanical Garden Edinburgh 40 (3): 553 (1983).
Pileo 15-37 mm, sottile, fragile, all’inizio conico-campanulato, poi convesso, con o senza un largo umbone ottuso negli esemplari maturi, margine involuto, infine reflesso, lungamente striato per trasparenza fino alla zona discale, superficie liscia, igrofana, a tempo umido subviscida, traslucida, di un bel colore aranciato lucente, ocra-aranciato, pallidescente per disidratazione a partire dal margine, infine interamente biancastra con tenui sfumature ocracee negli esemplari maturi e ben disidratati; resti velari presenti soltanto nei giovani esemplari sotto forma di minute fioccosità biancastre facilmente detersili.
Lamelle arrotondato-annesse, dapprima moderatamente fitte, poi un po’ più spaziate, ventricose, larghe fino a 4-5 mm, intercalate da 3-4 lamellule, dapprima bianche, poi bianco-grigiastre, infine ocracee, filo finemente crenulato, biancastro.
Stipite 35-60 x 2-5 mm, cilindrico, leggermente allargato alla base, slanciato, fistoloso, superficie inizialmente bianca e ricoperta da una fine pruina concolore, in seguito leggermente ingrigente, fino a bruno-ocracea a partire dalla base per imbibizione.
Carne sottile, fibrosetta nel gambo, biancastra, inodora, sapore mite.
Sporata non ottenuta.
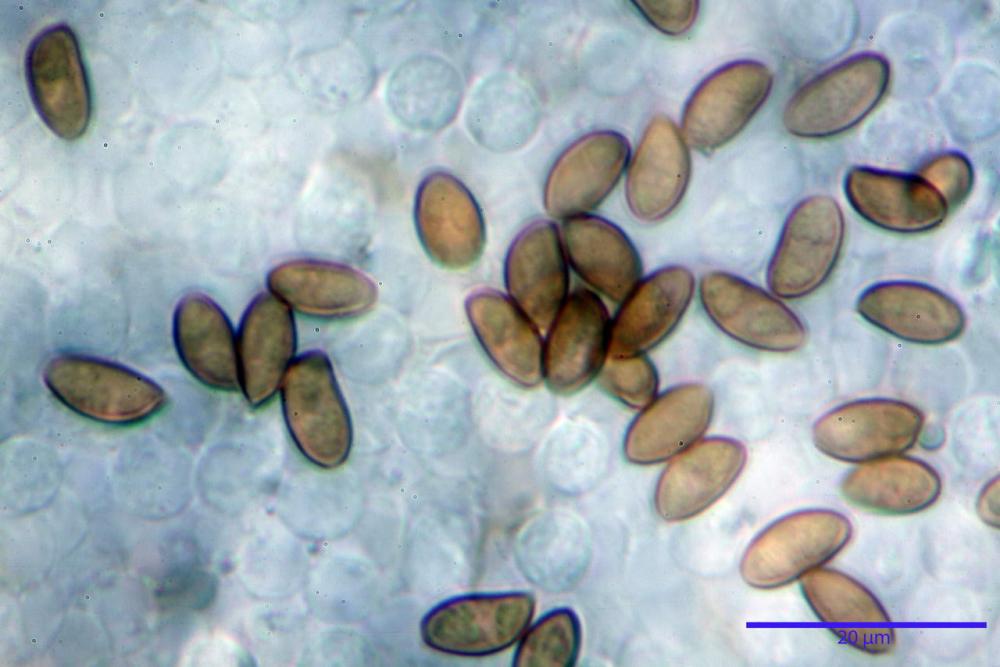
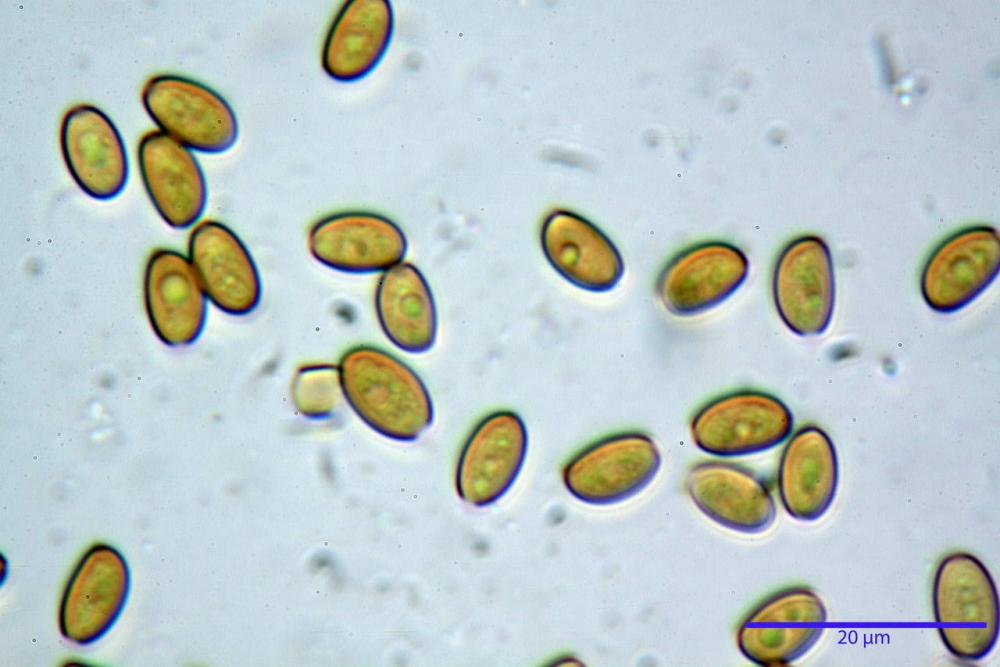
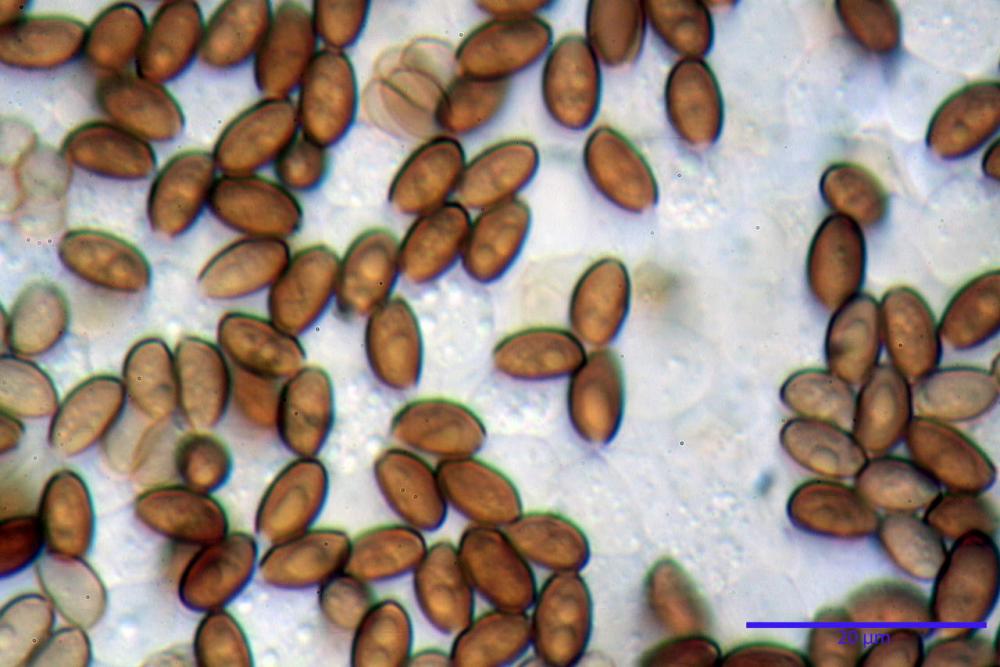
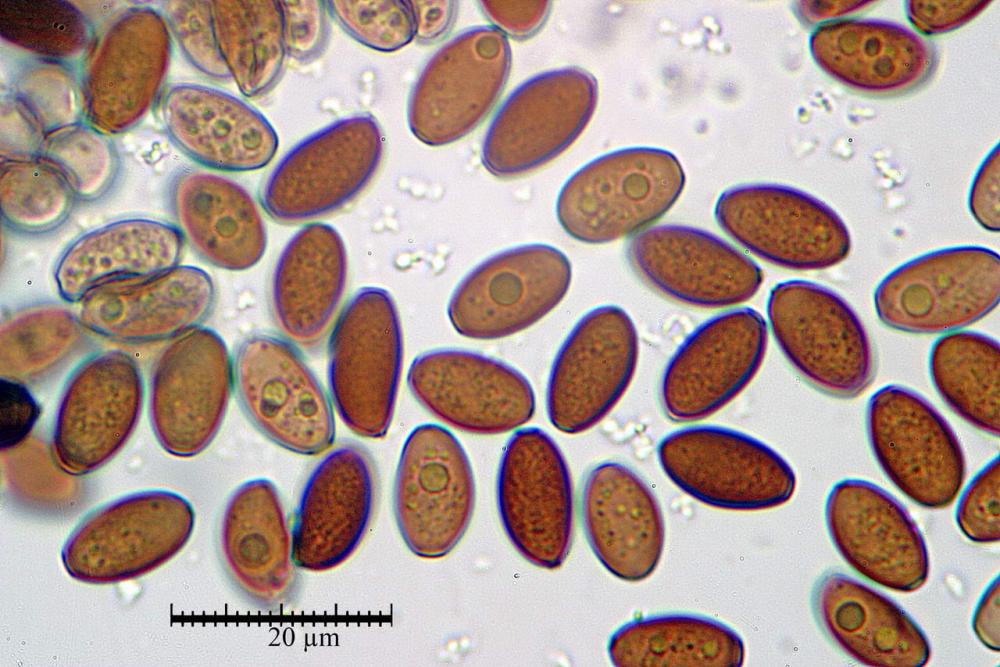
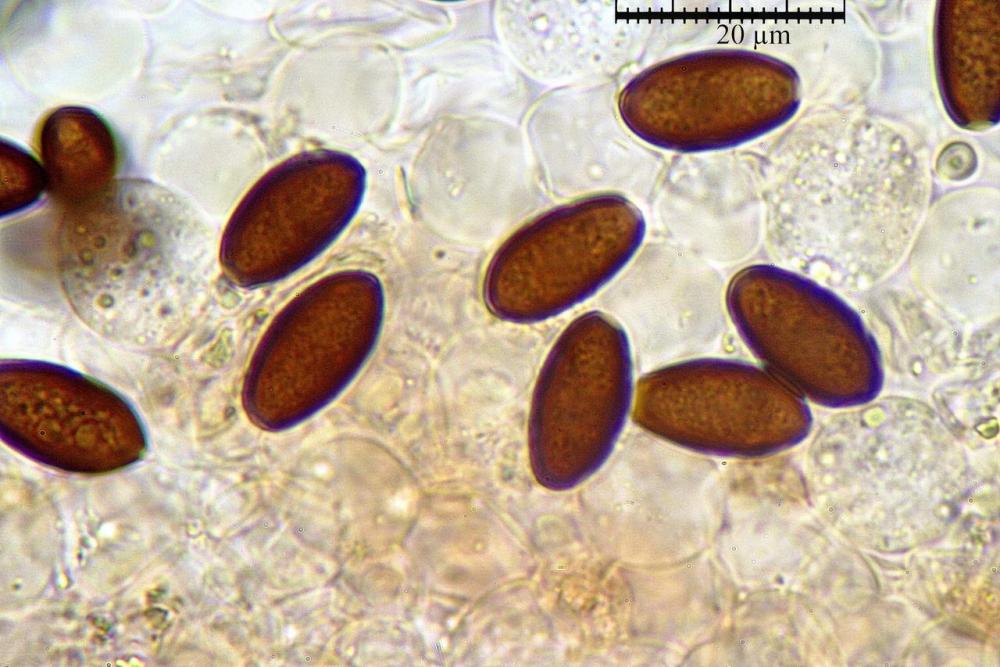
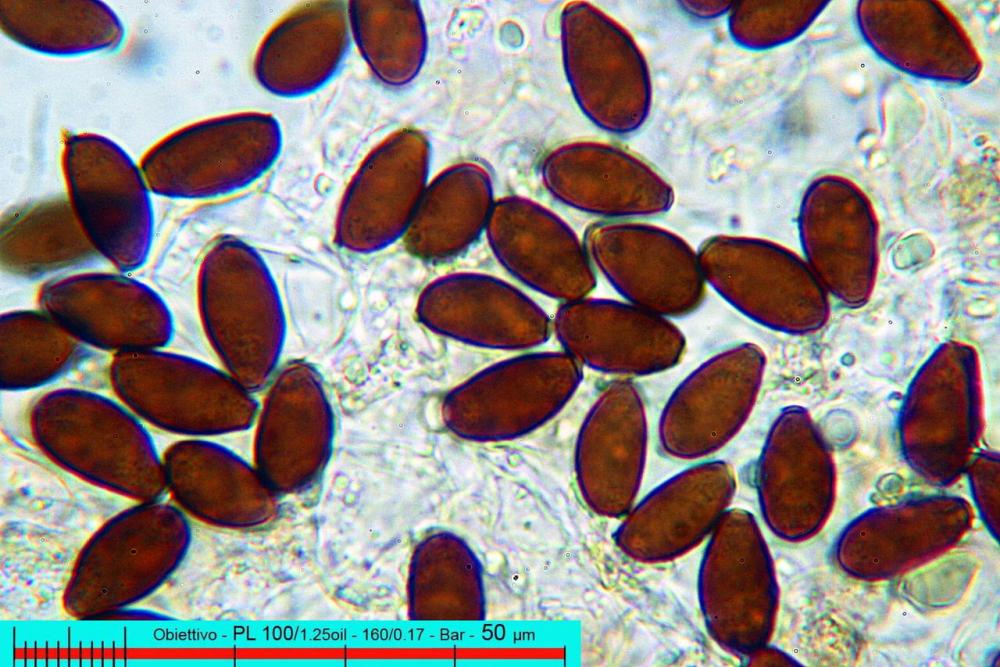
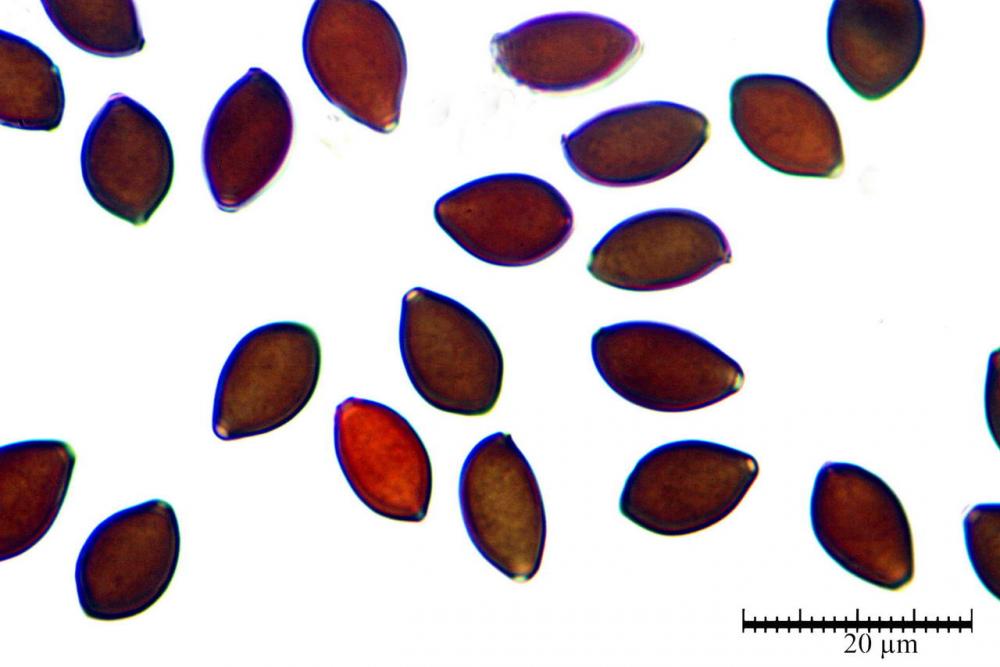
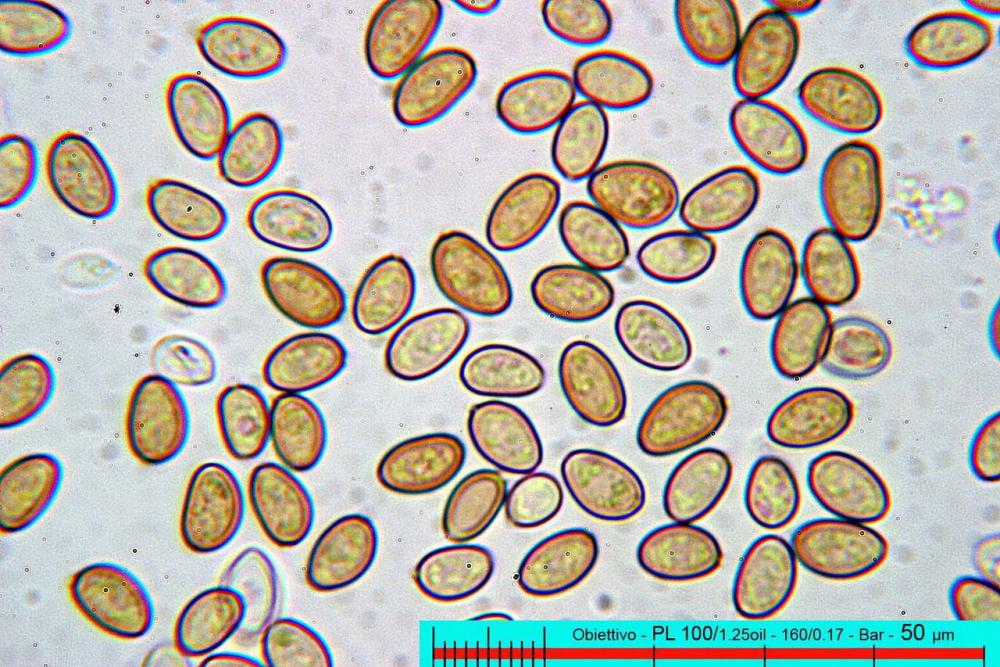
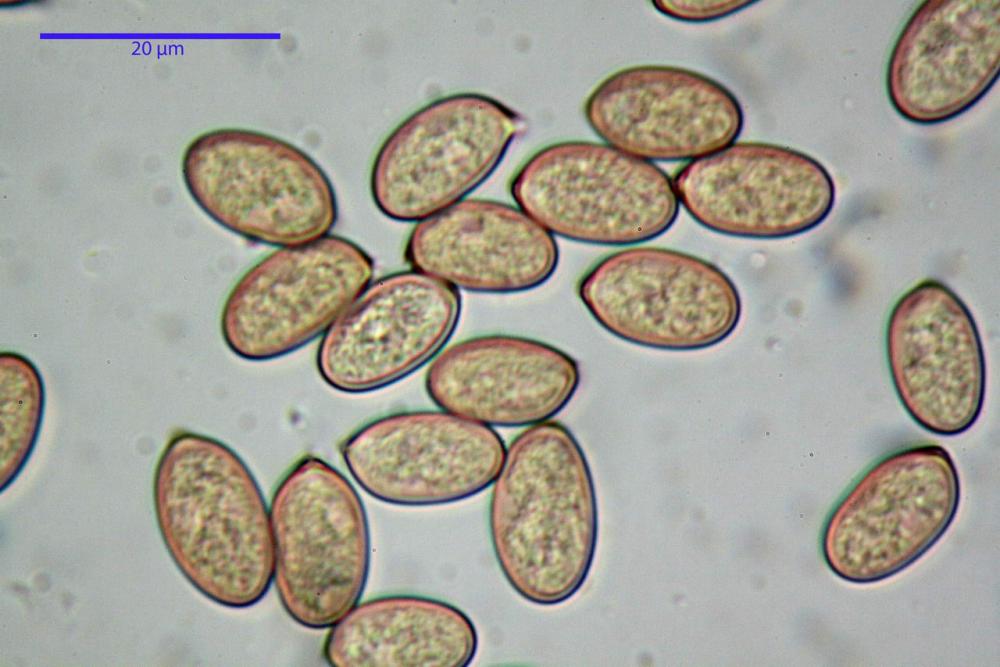
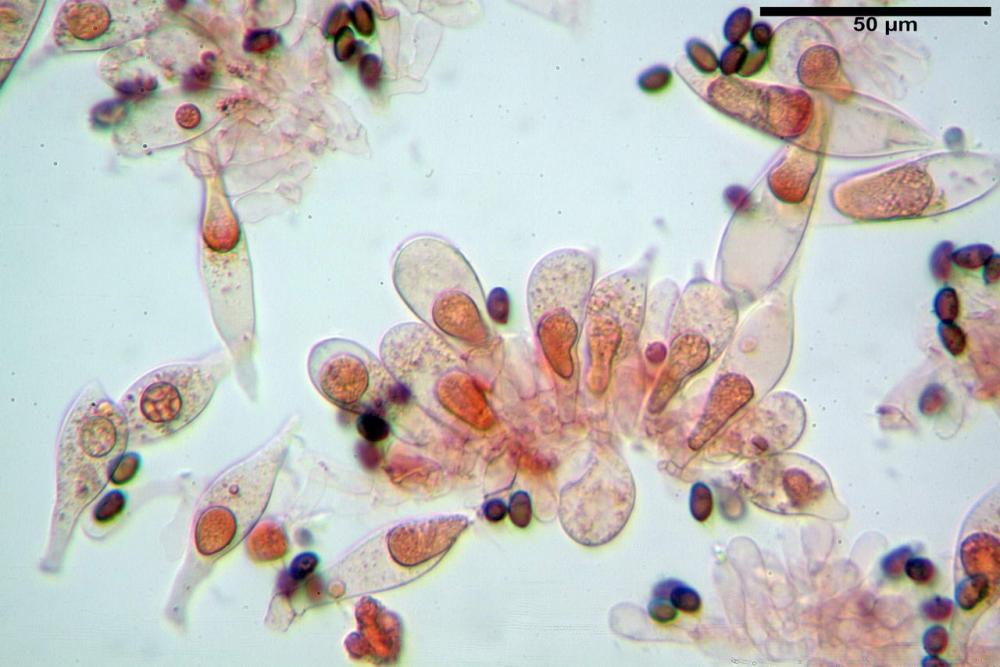
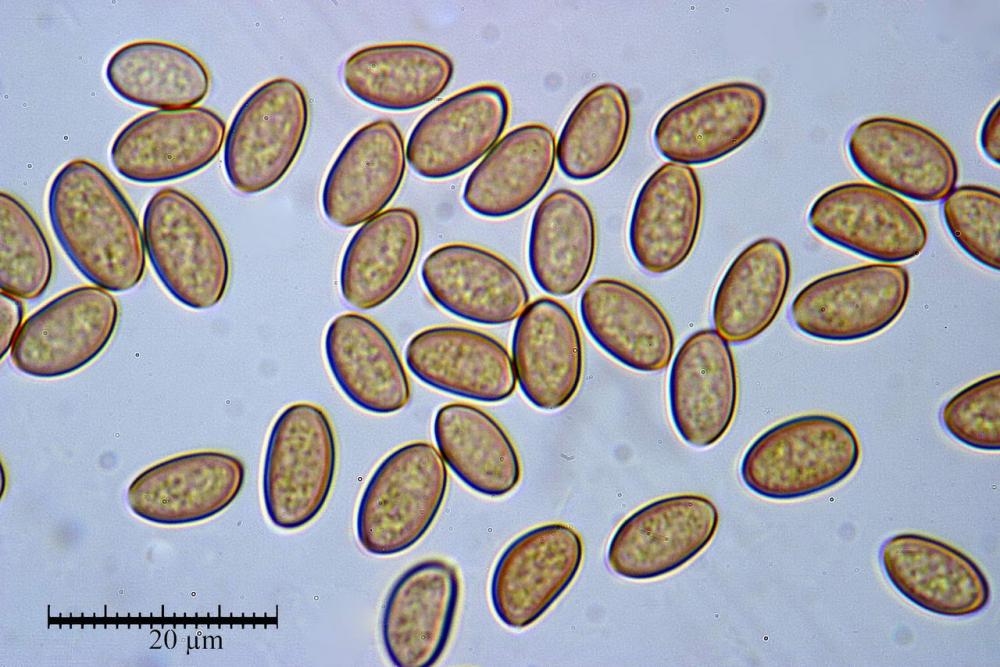
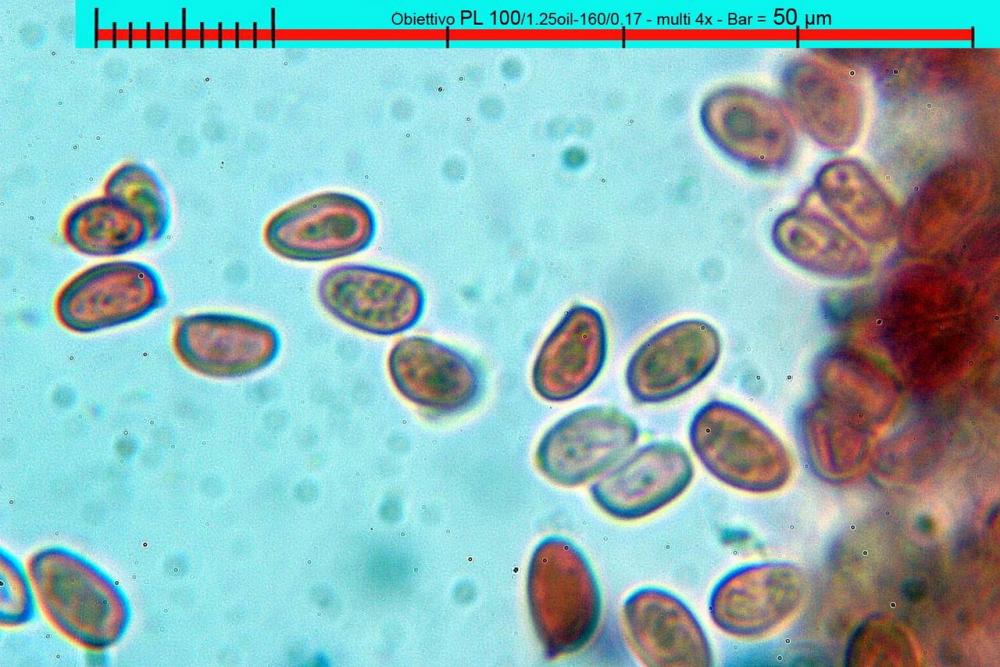
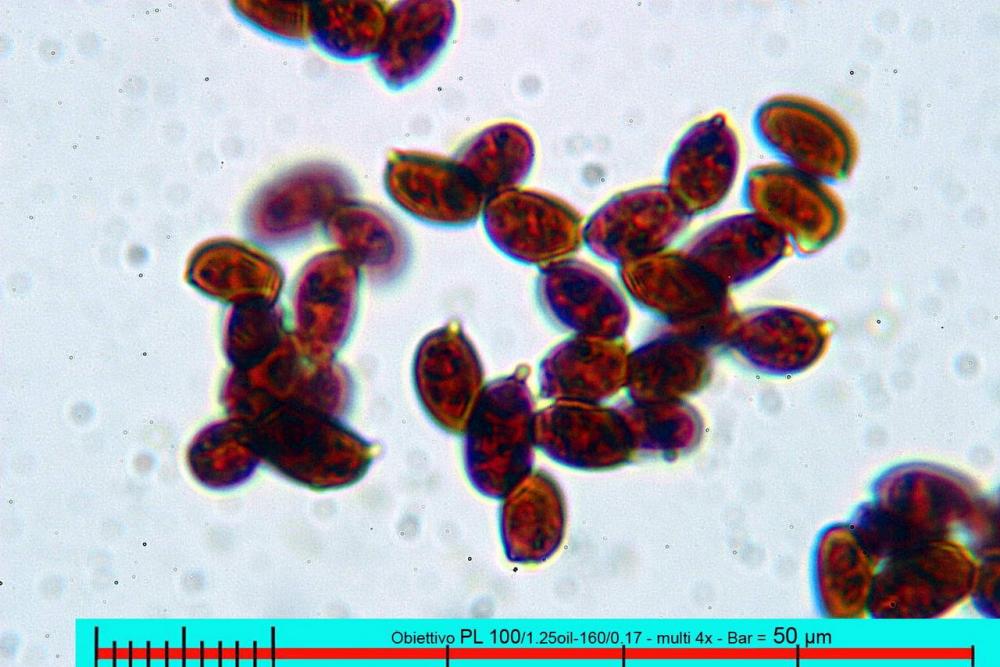
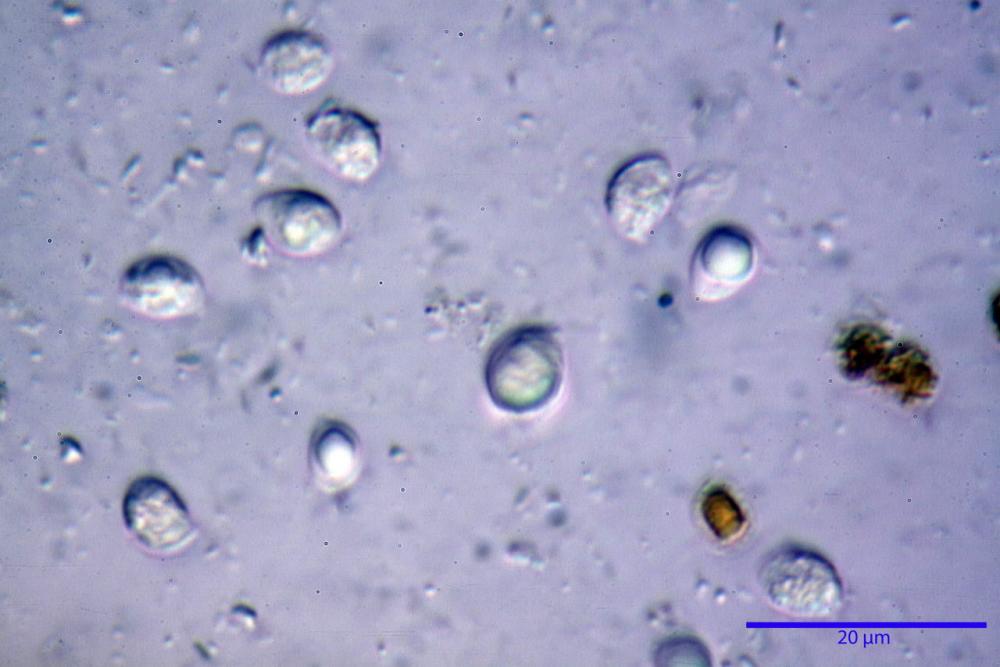
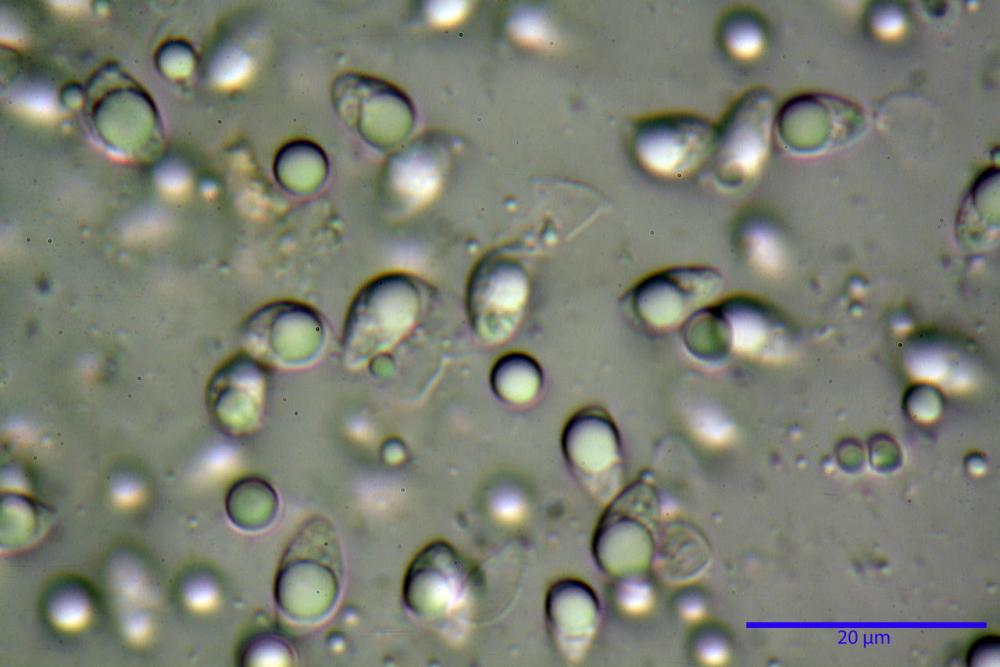
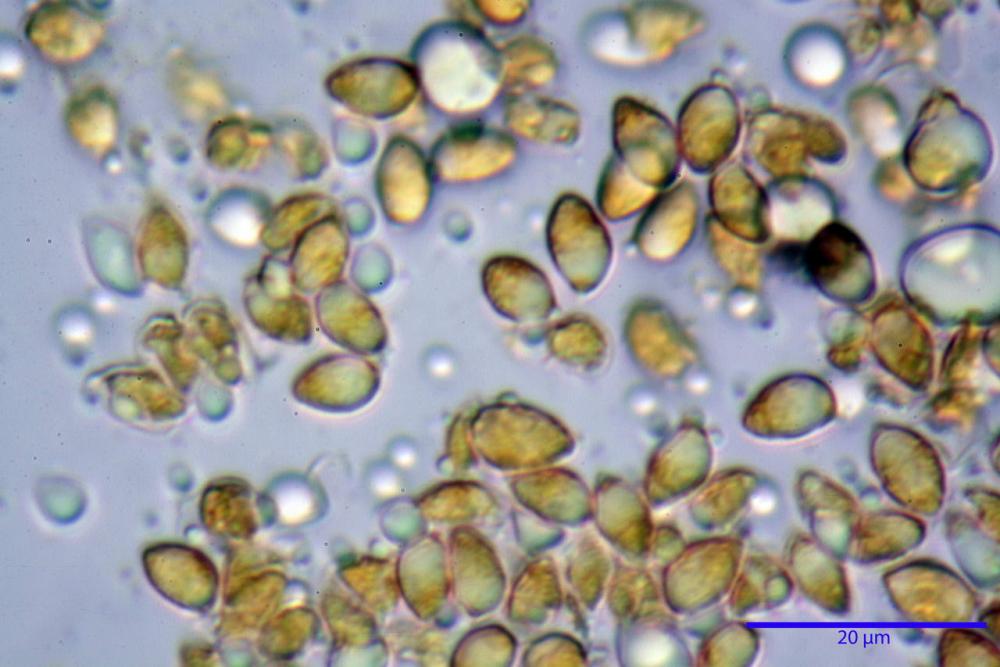
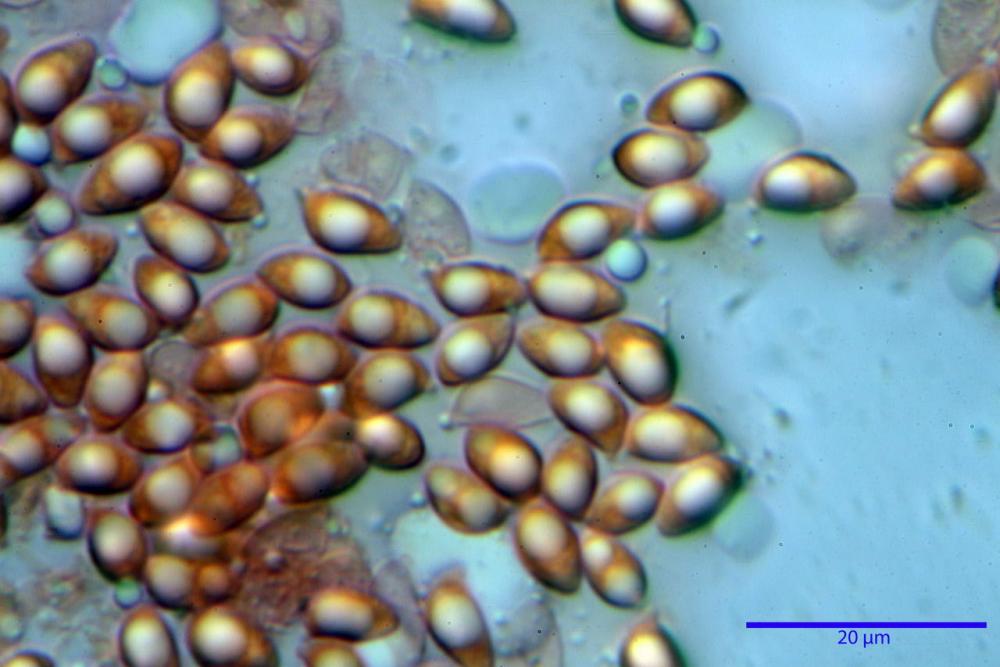
Spore (7,7)8,4-10,1(11,3) x (4,4)4,6 -5,2(5,5) µm, in media 9,3 x 4.9 µm, Q. = (1,5)1,7 - 2,0(2,2), Q.m. = 1.9, Vol.m. = 119,2 µm³, da ellissoidi con profilo adassiale appiattito fino a subamigdaliformi in proiezione laterale, ovoidi in proiezione ventrale, apparentemente lisce, a forti ingrandimenti dimostrano una leggera punteggiatura, poro germinativo largo 0,75-1,20 µm, alle volte poco visibile sottoforma di callus.
Basidi 26,9-38,9 x 7,2- 9,8 µm, in media 30,9 x 8,3 µm, tetrasporici, clavati, non notati basidi bisporici.
Trama imeniale composta da ife inflate, con diametro di 5,5-24,5 µm.
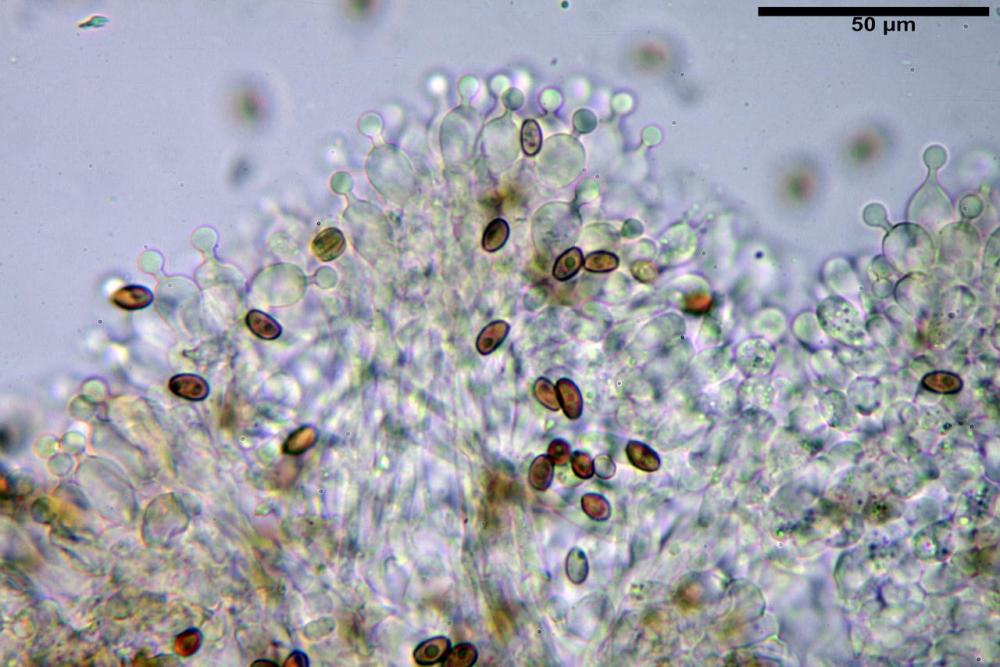
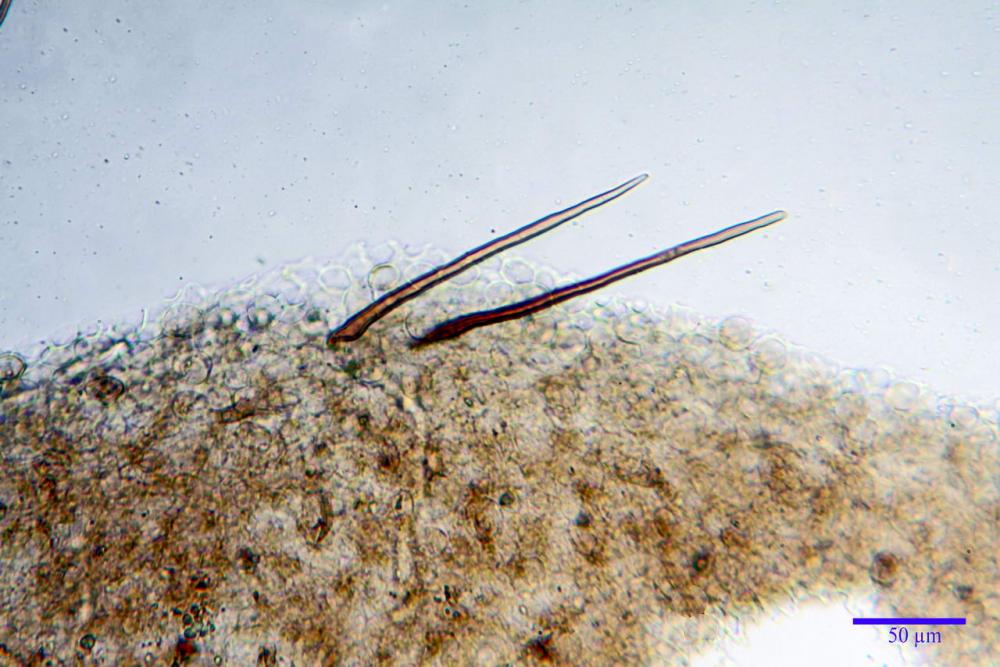
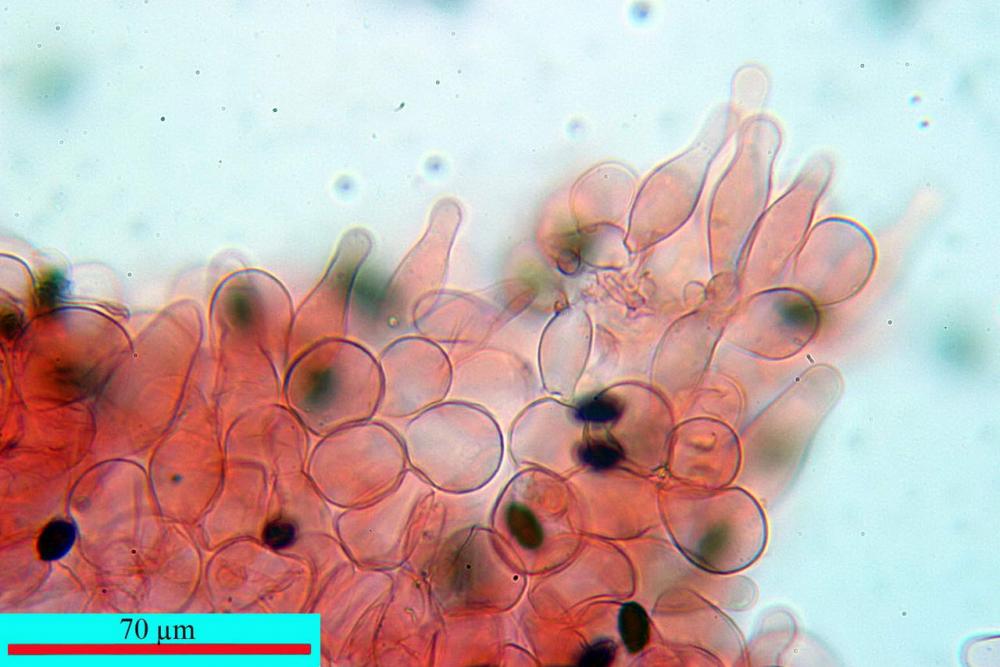
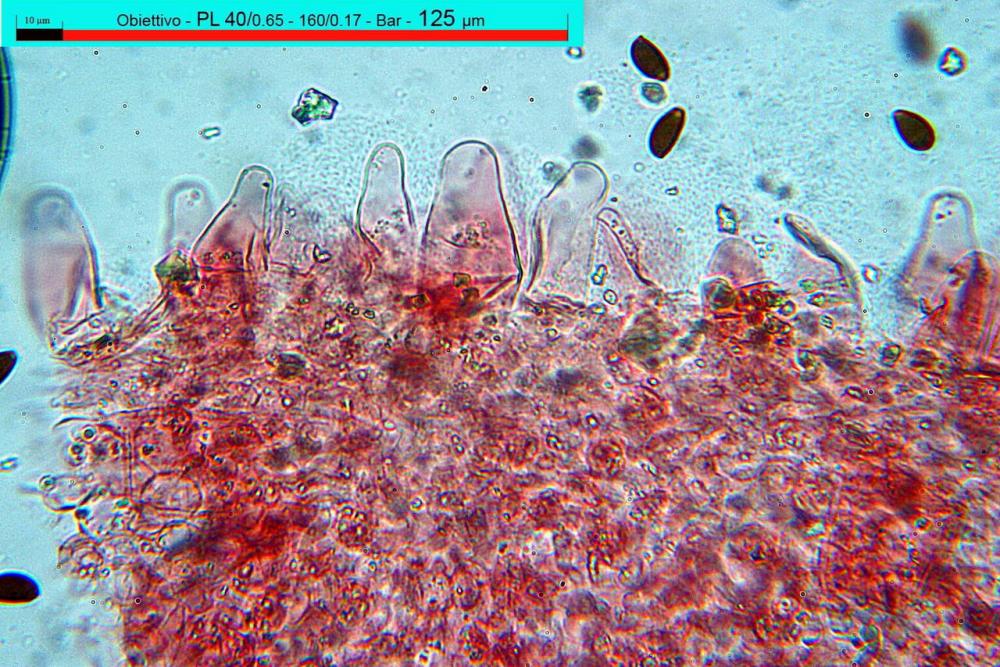
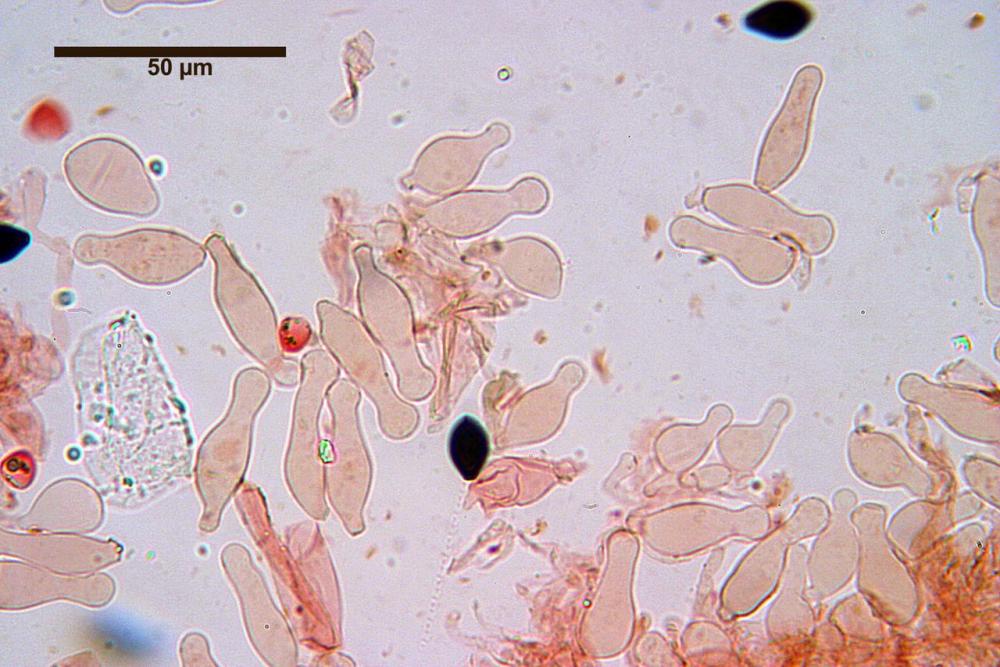
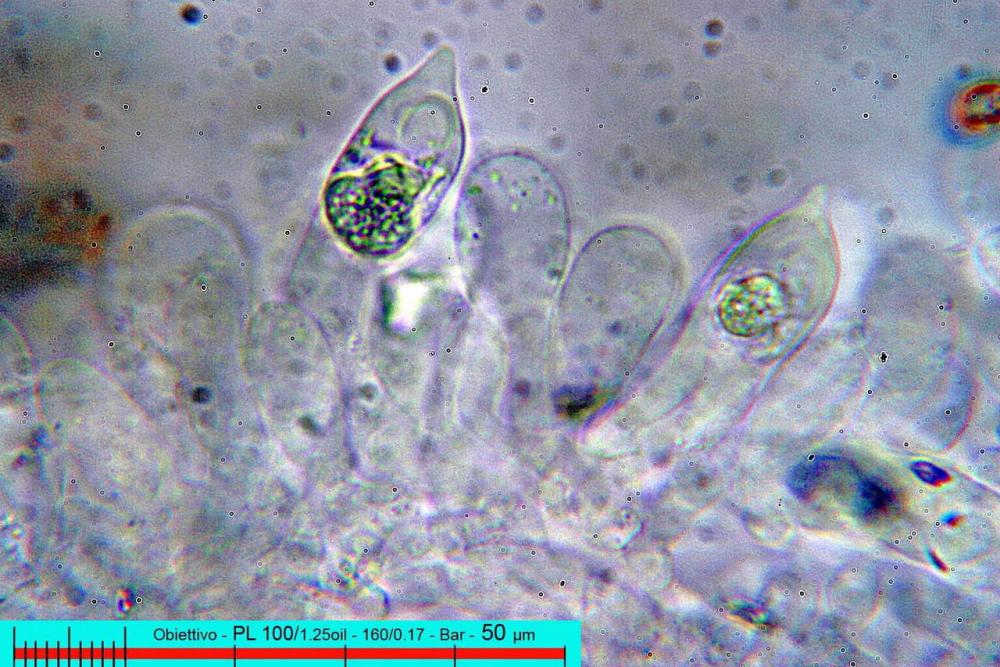
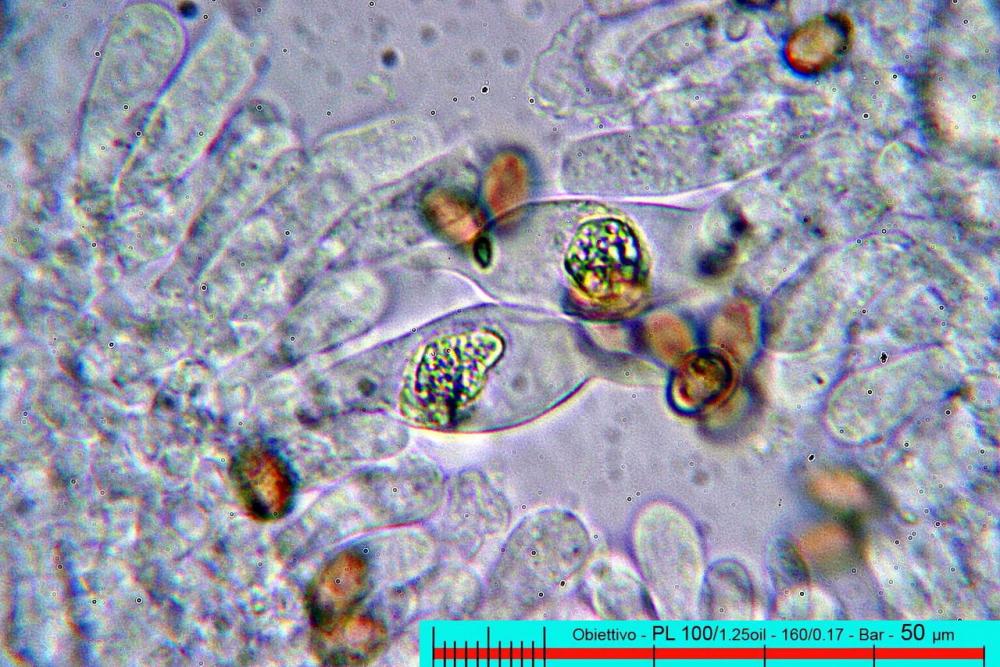
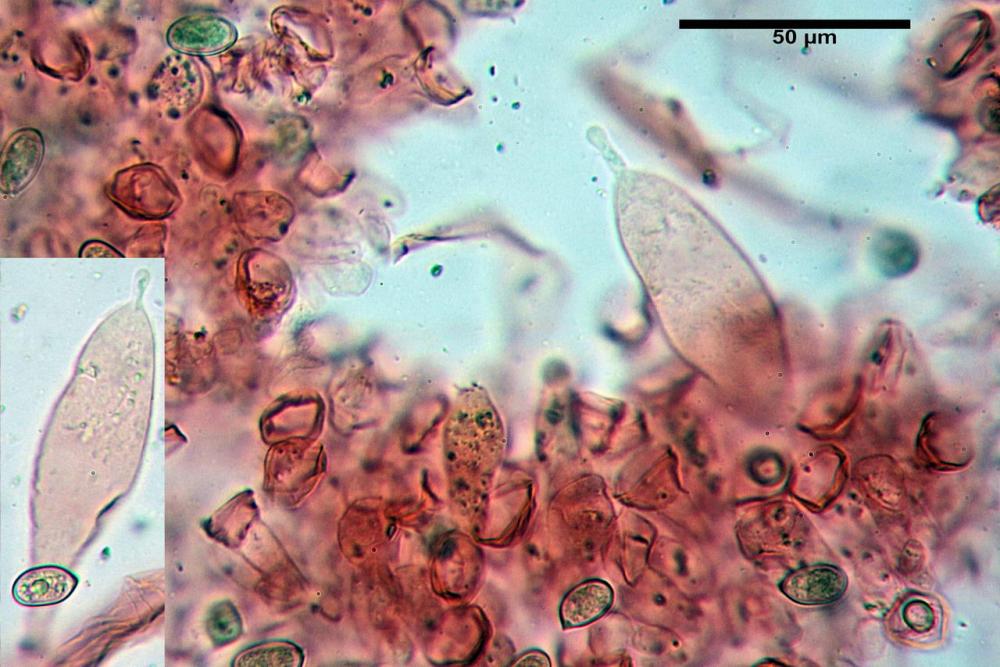
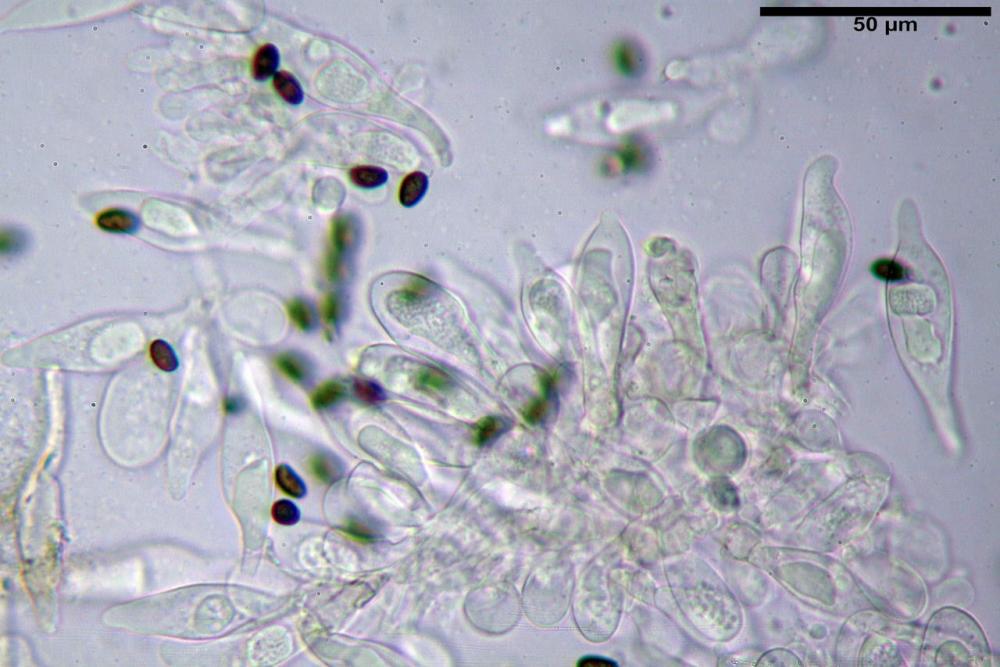
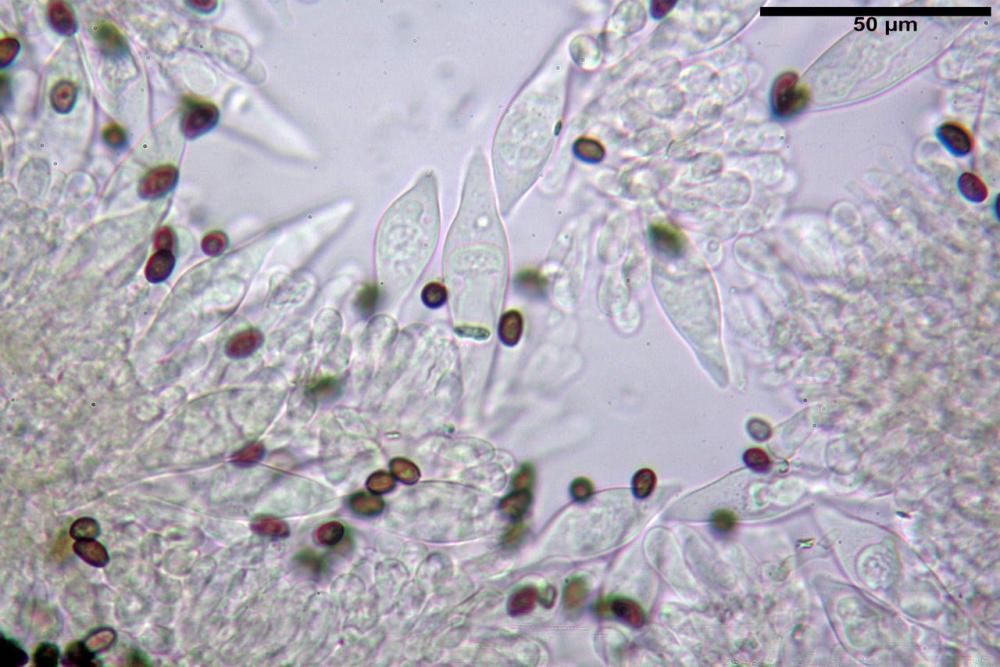
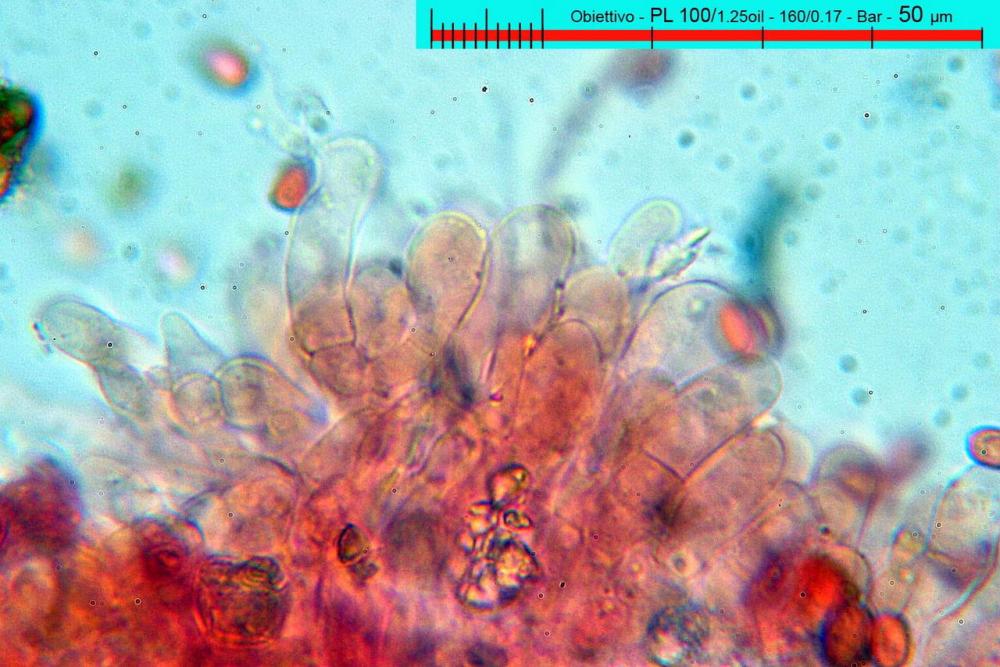
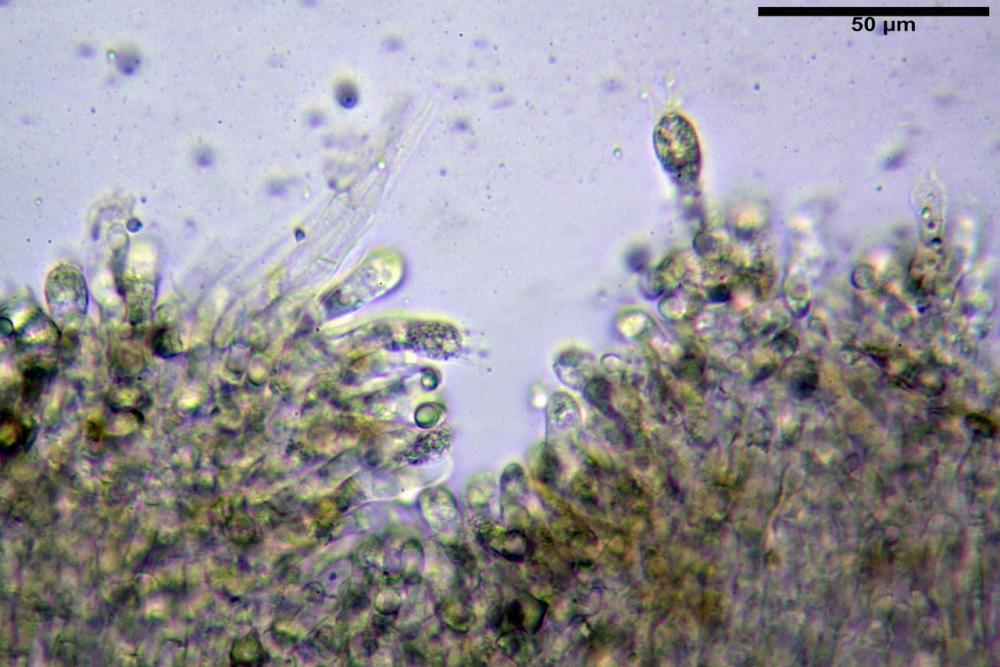
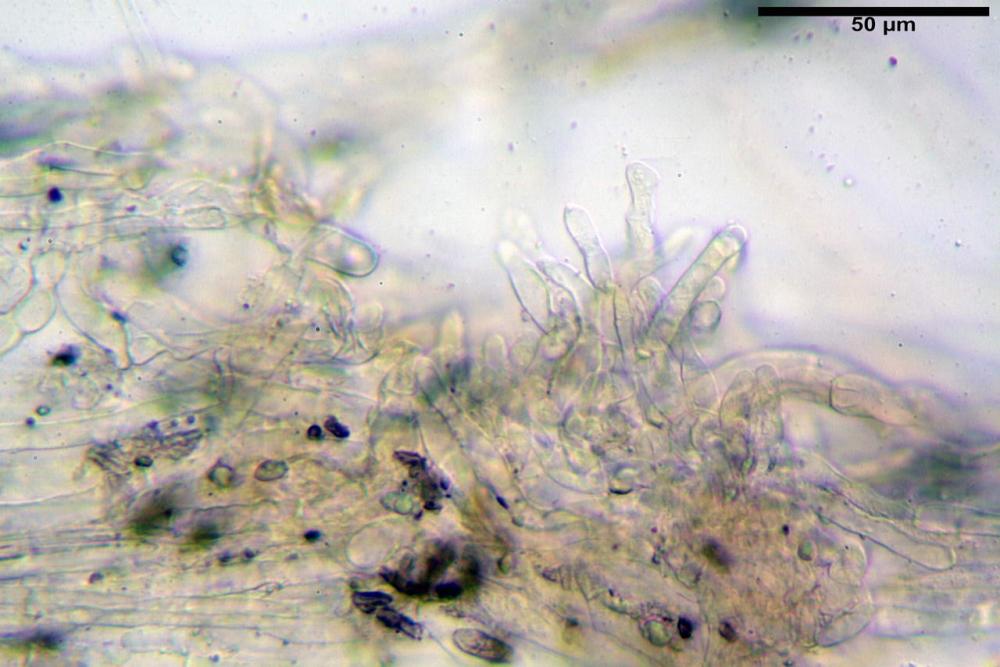
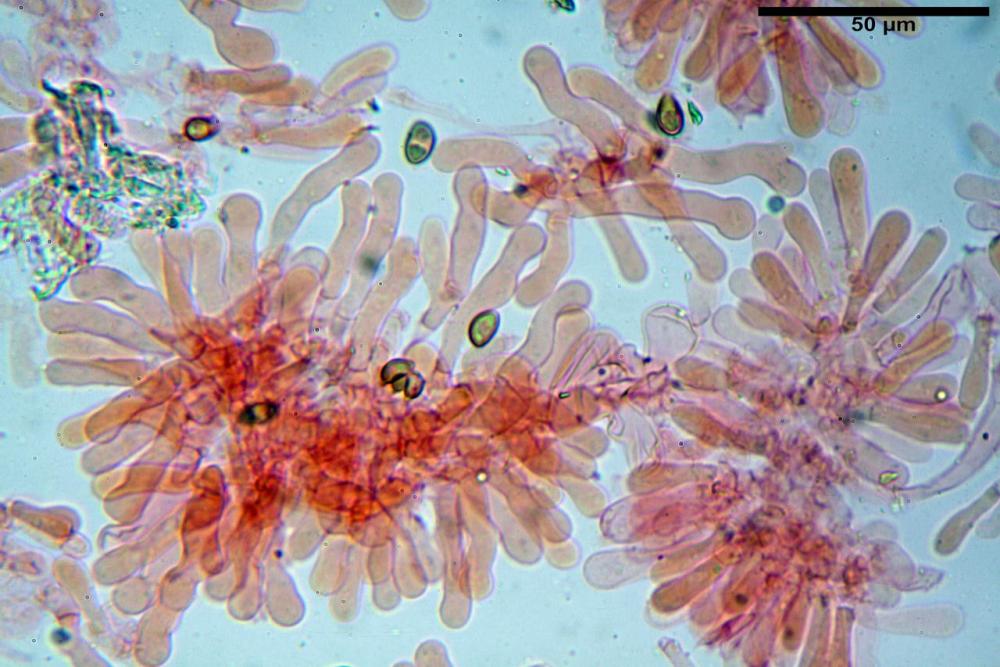
Cheilocistidi (17,4)30,4-39,0(44,8) x (6,6)6,7-10,7(12,0) µm, in media 35,4 x 8,5 µm, cilindrici e flessuosi, alle volte con apice allargato (Opuntia), subfusiformi, clavati e in questo caso molto spesso settati, densamente disposti su tutto il filo lamellare tanto da renderlo sterile.
Pleurocistidi assenti.
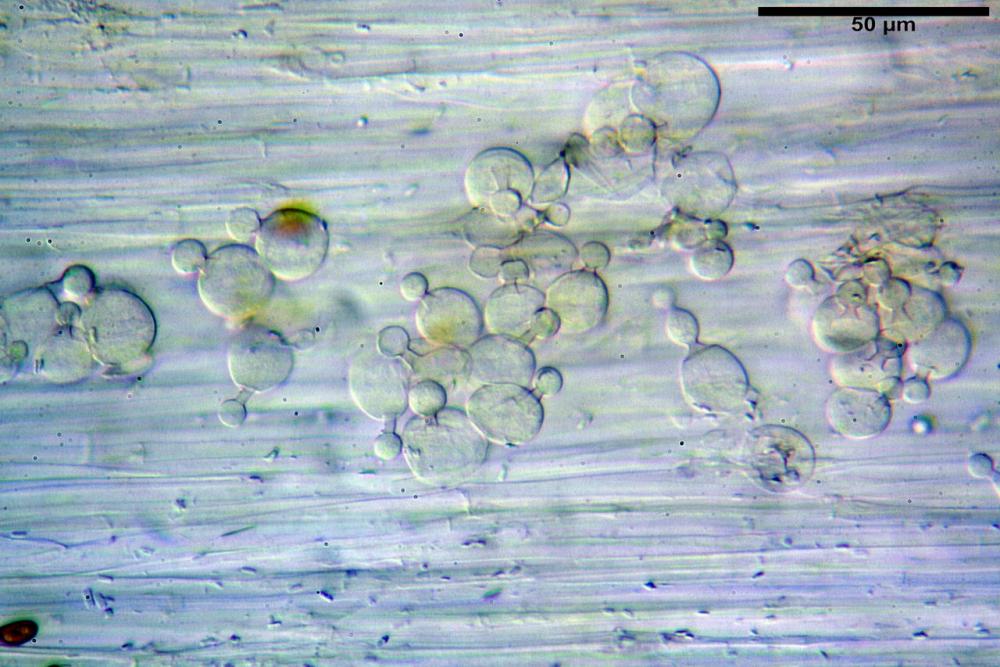
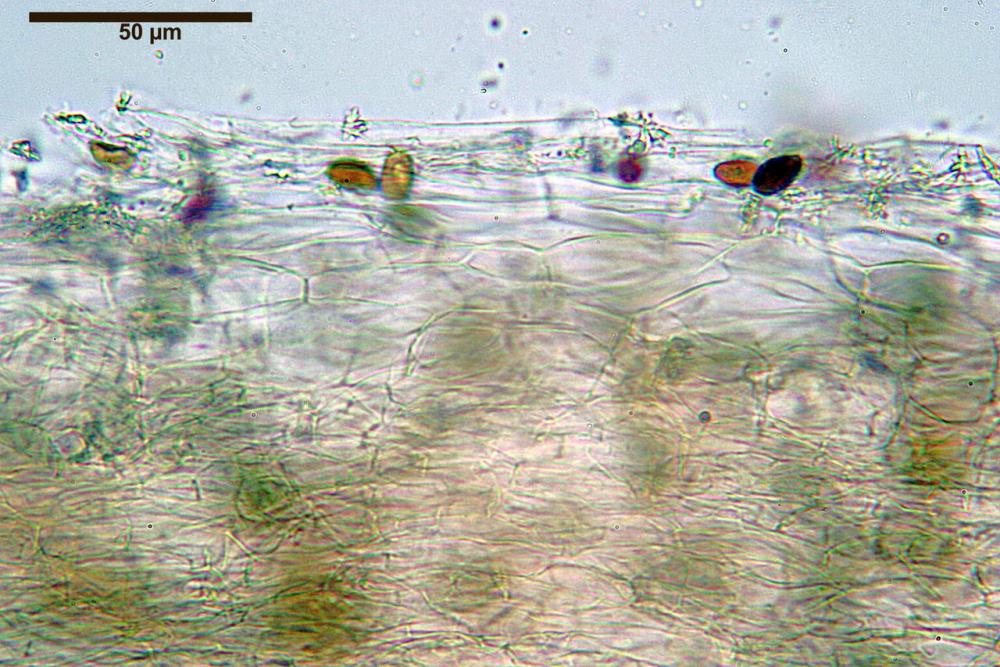
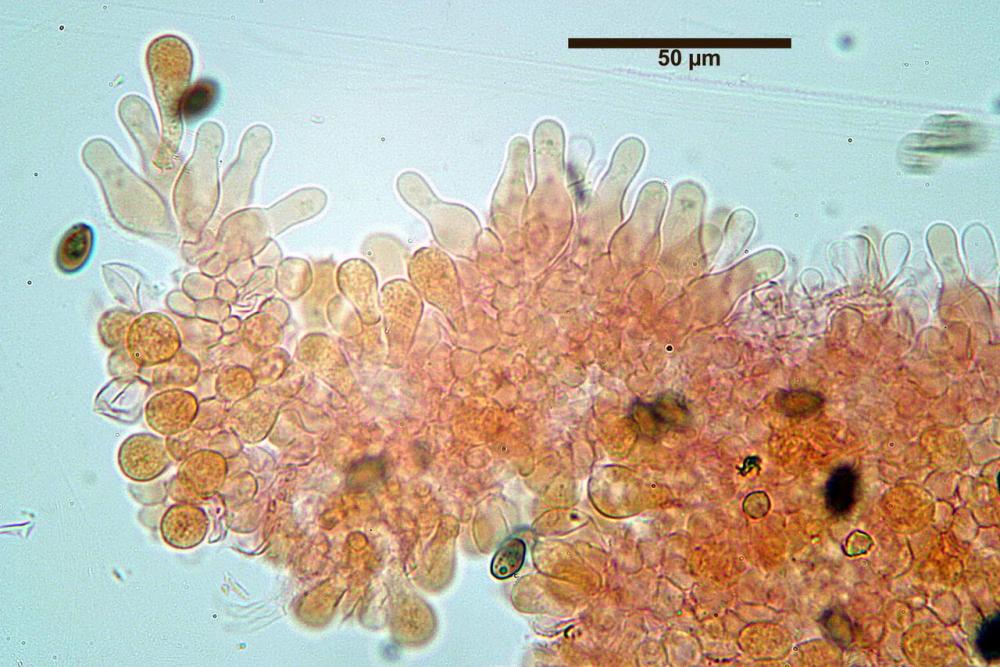
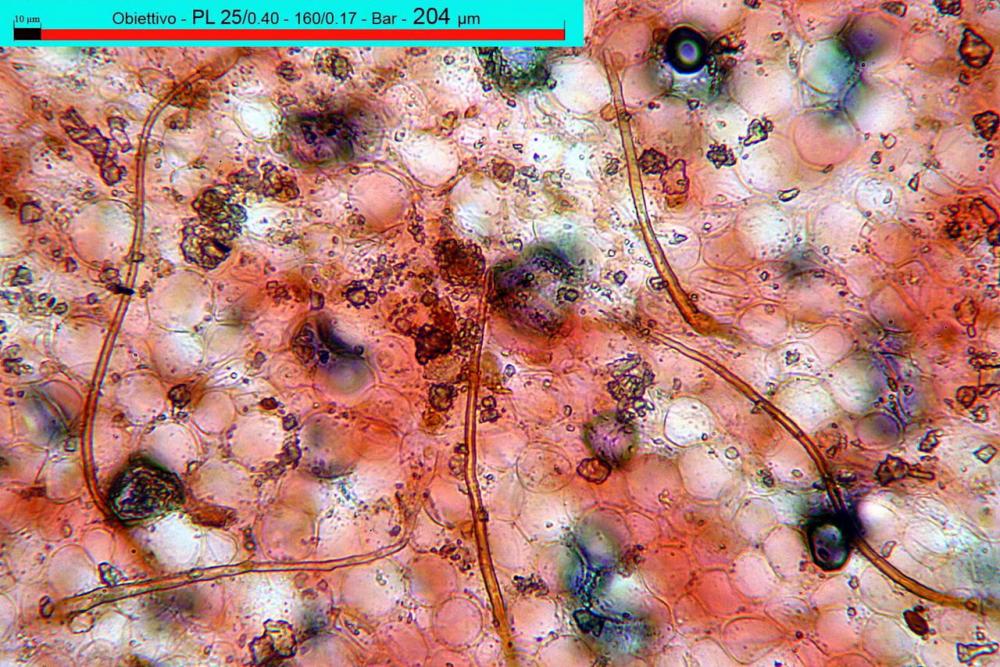
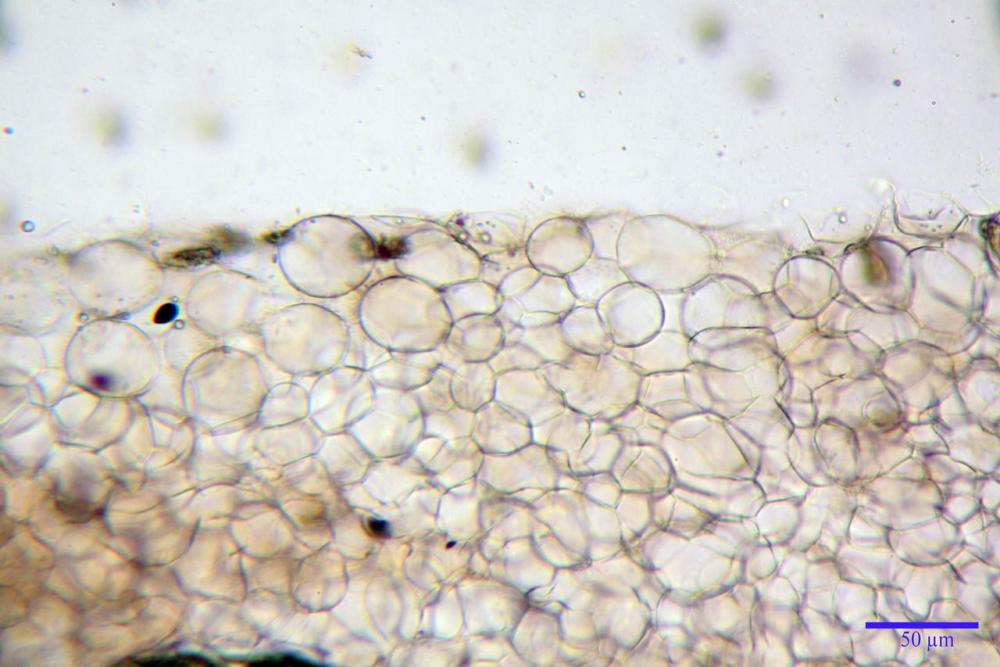
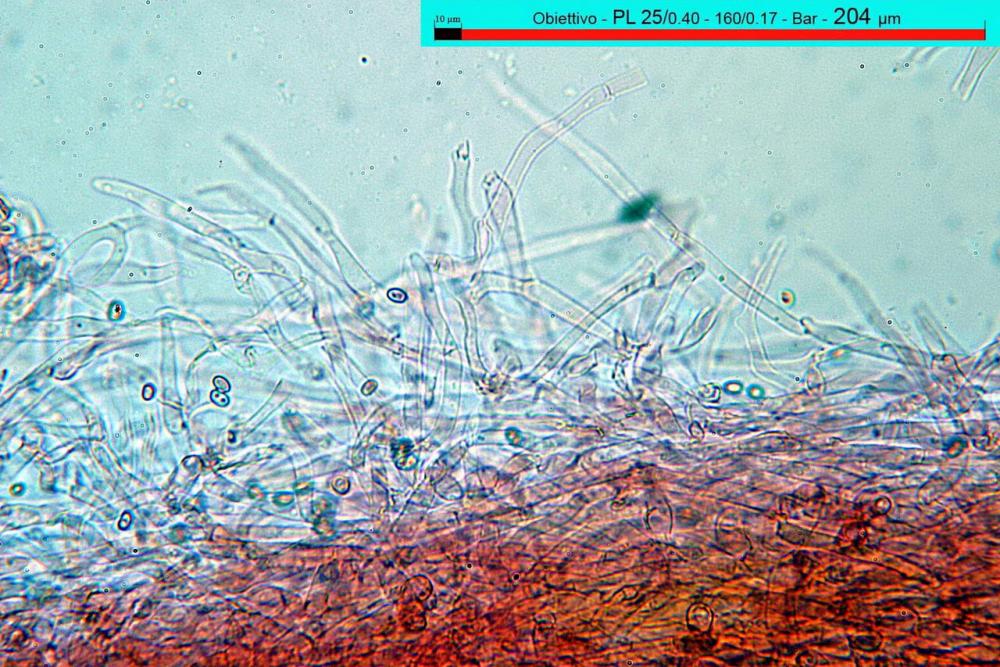
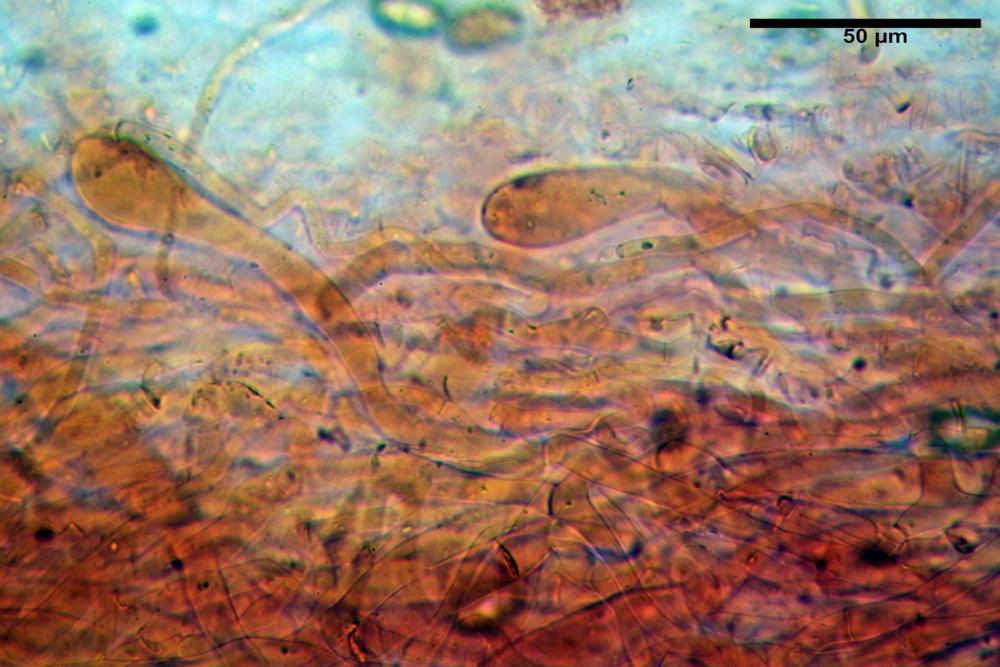
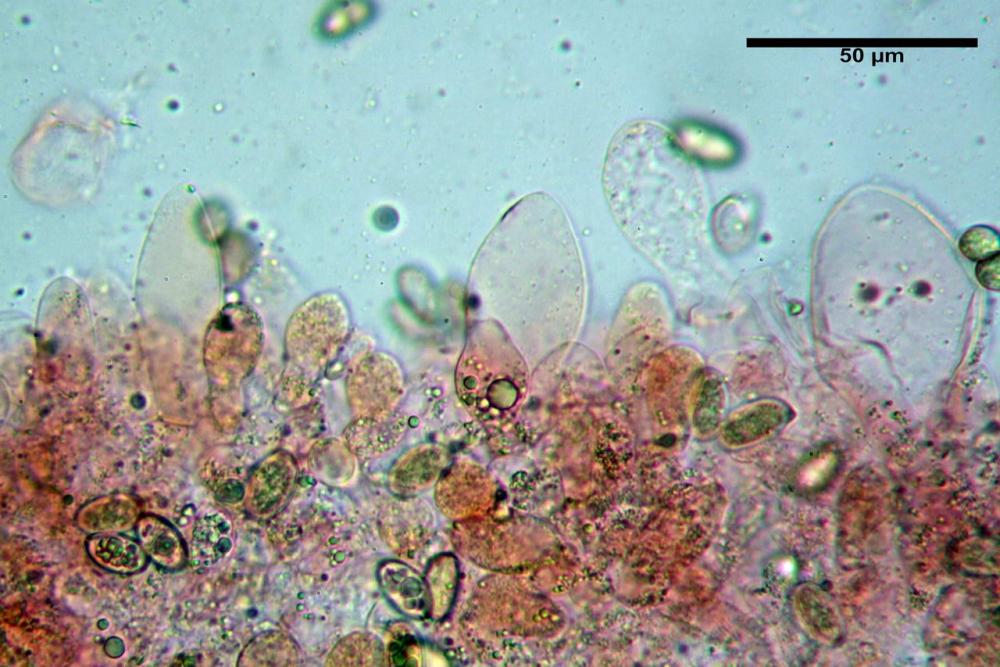
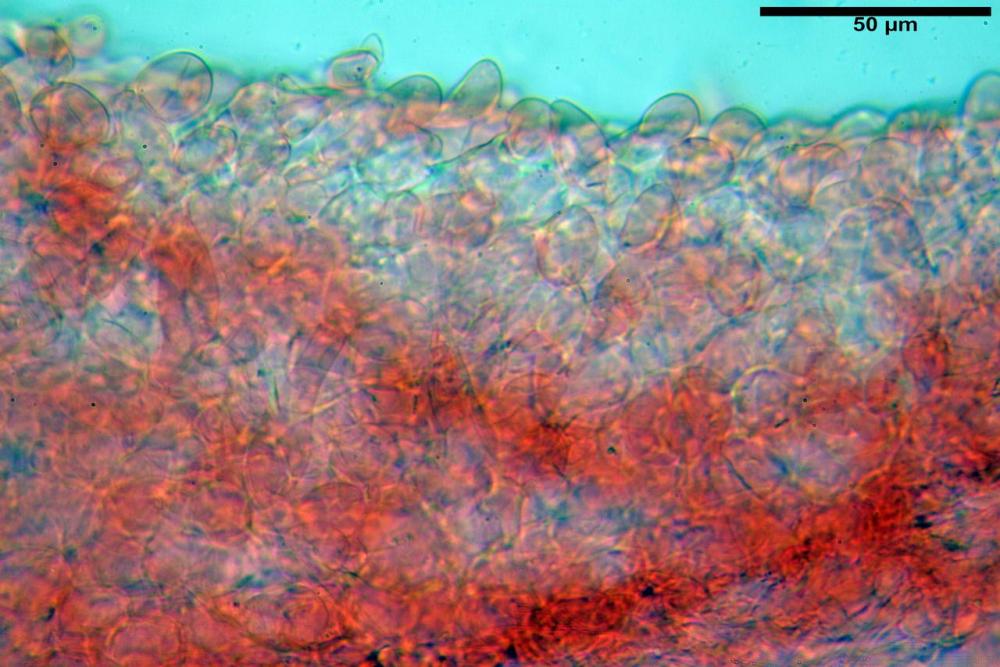
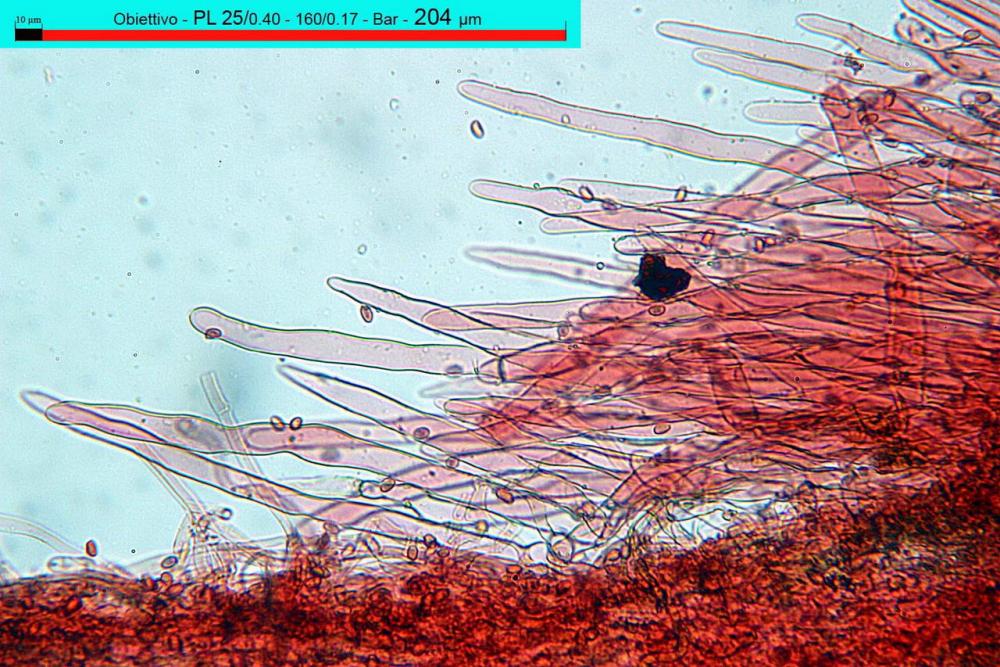
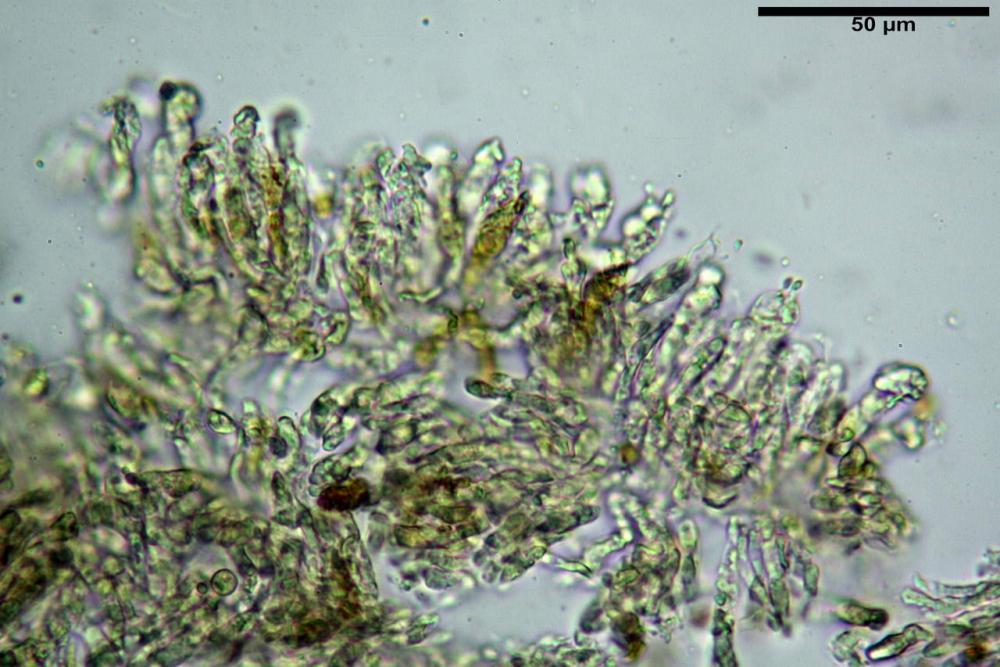
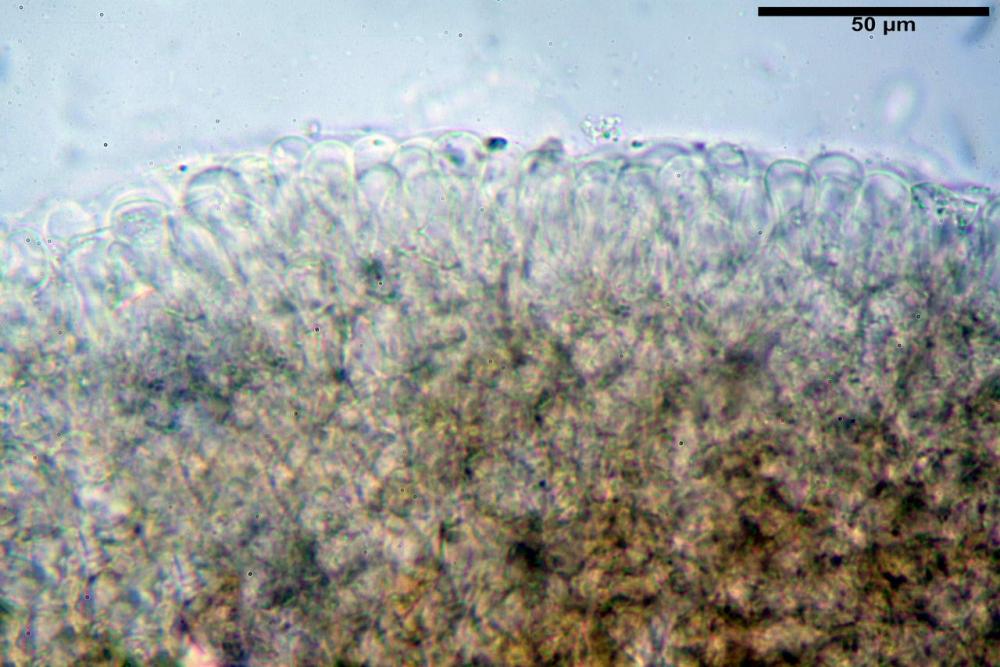
Pileipellis di tipo imeniderma, composta da cellule sferopeduncolate, clavate o piriformi, ialine, di dimensioni (33,5)34,3-51,9(56,4) x (10,4)11,4-17,1(17,3) µm, in media 43,2 x 14,0 µm.
Pileocistidi assenti.
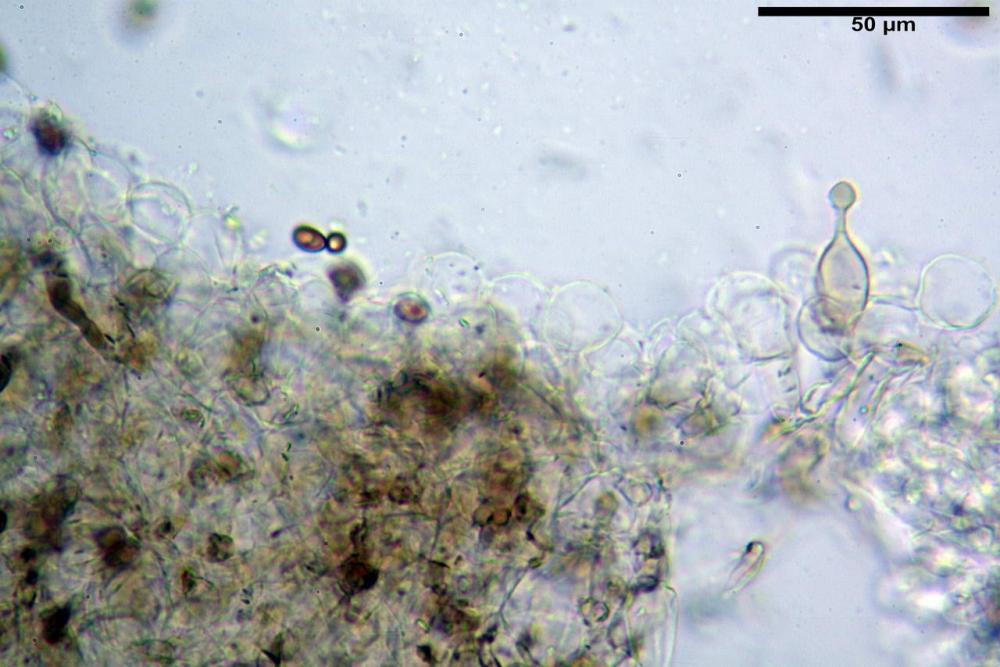
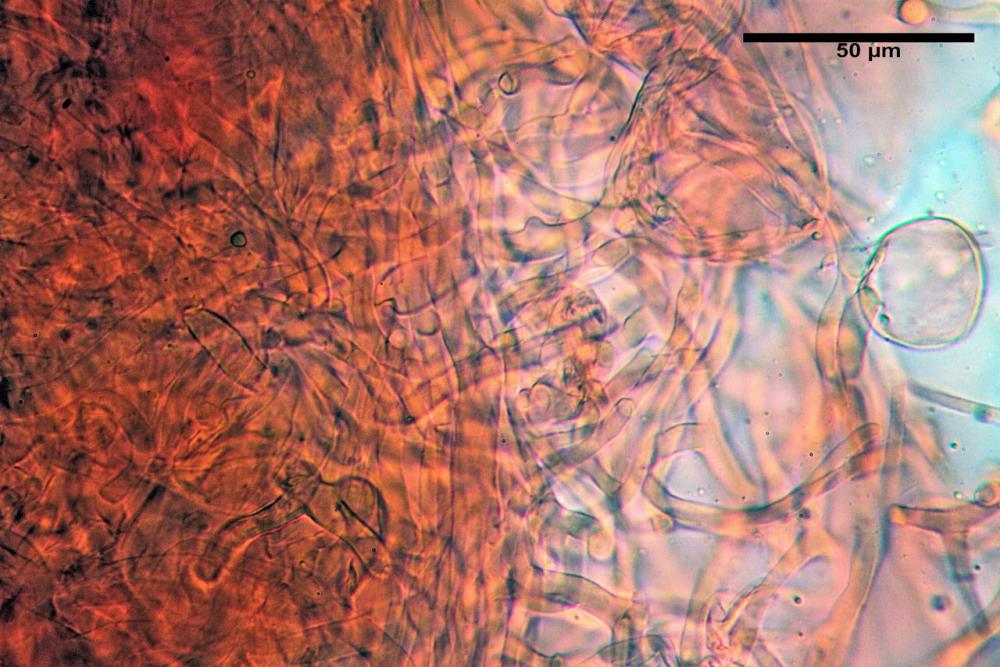
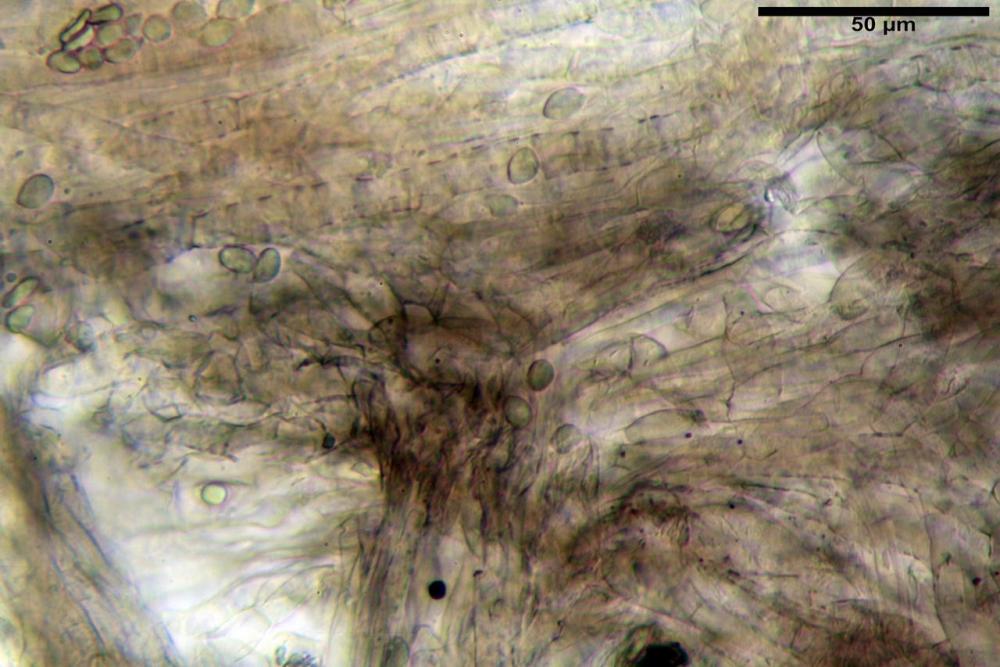
Caulopellis tipo cutis, formata da ife di diametro (3,7)4,0-7,6 (11,0) µm, cilindriche, settate, ialine.
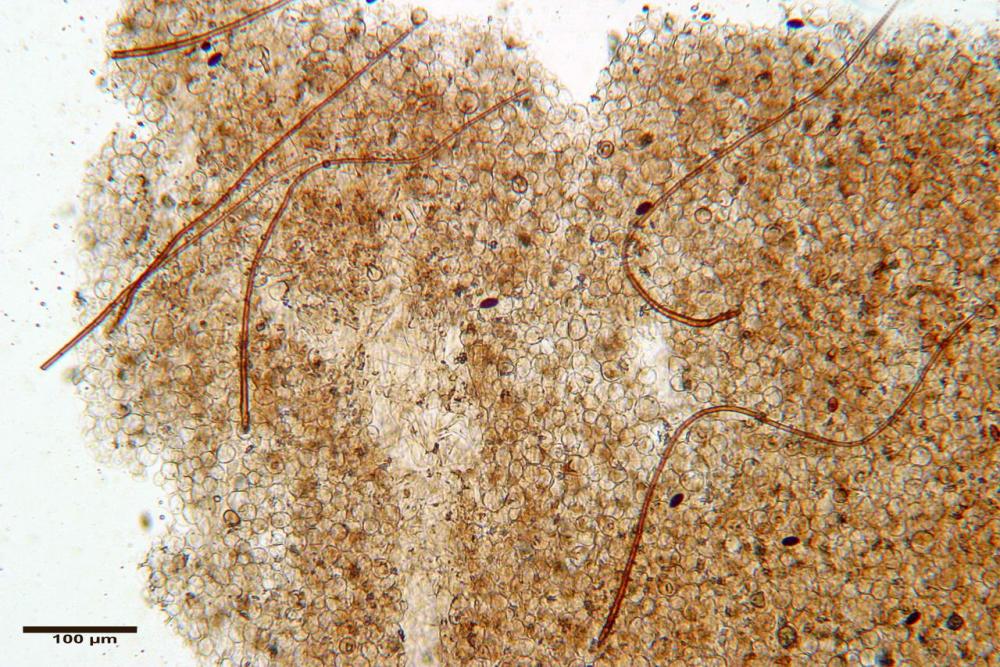
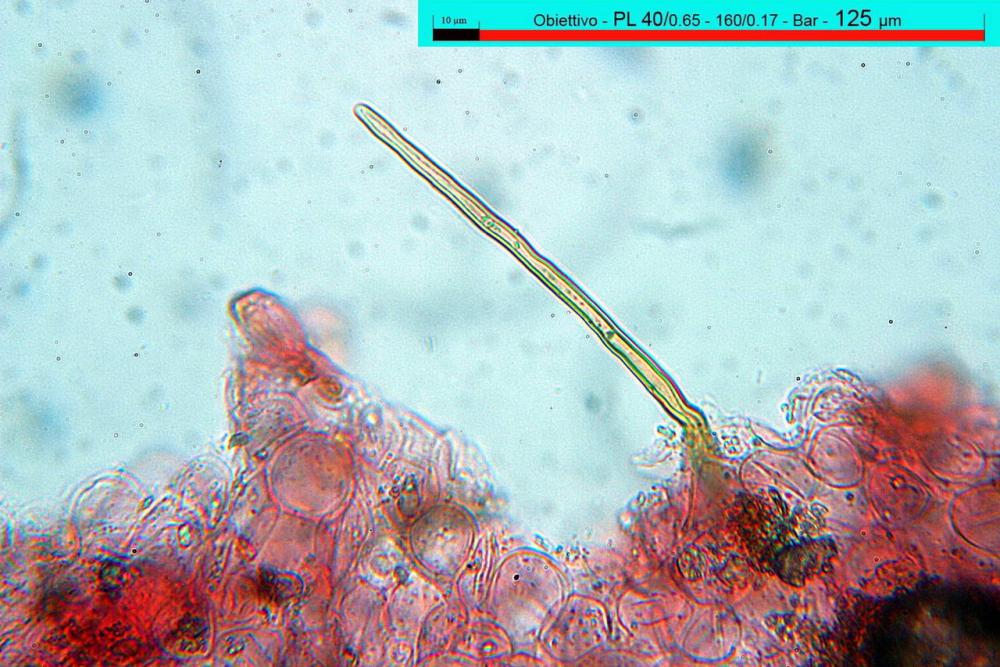
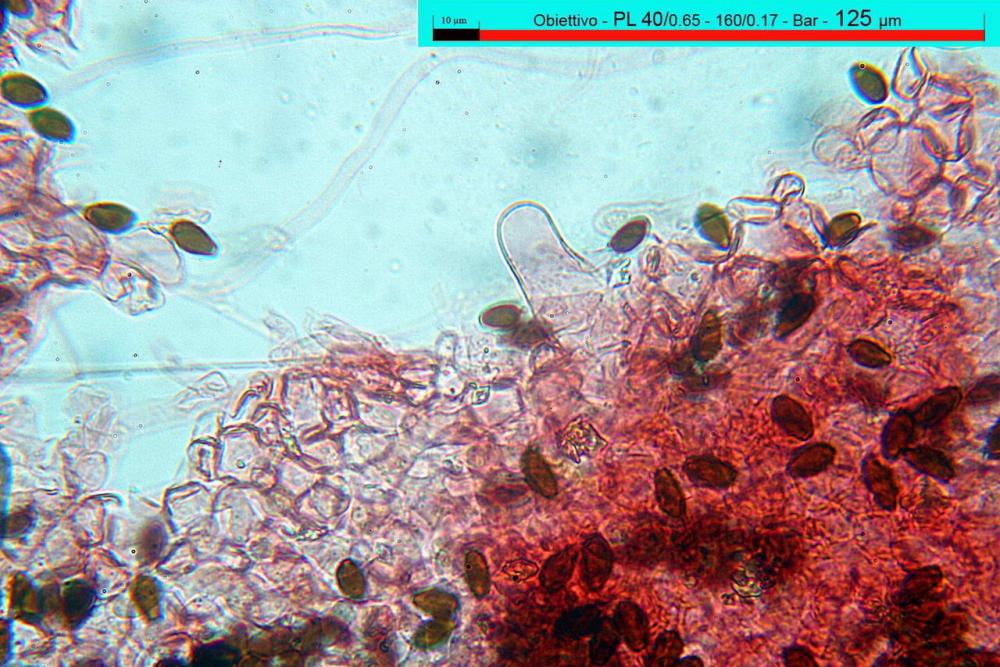
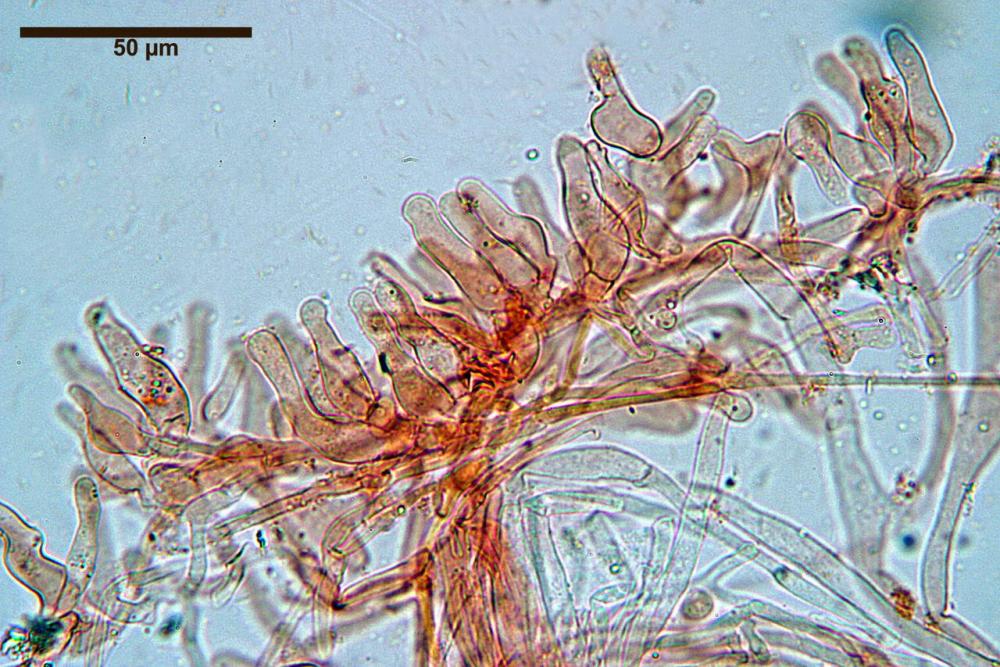
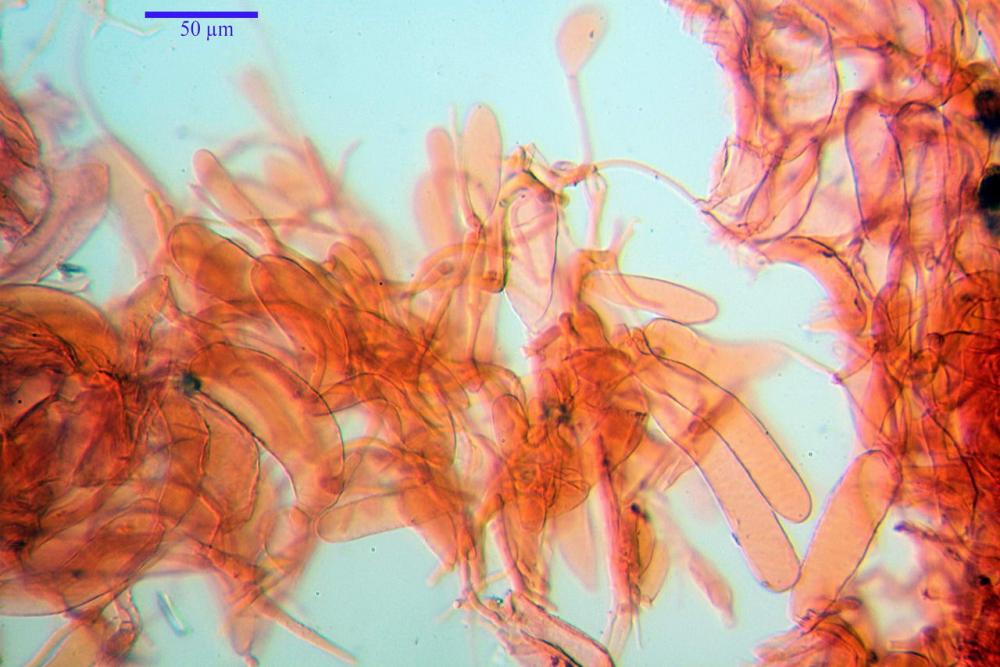
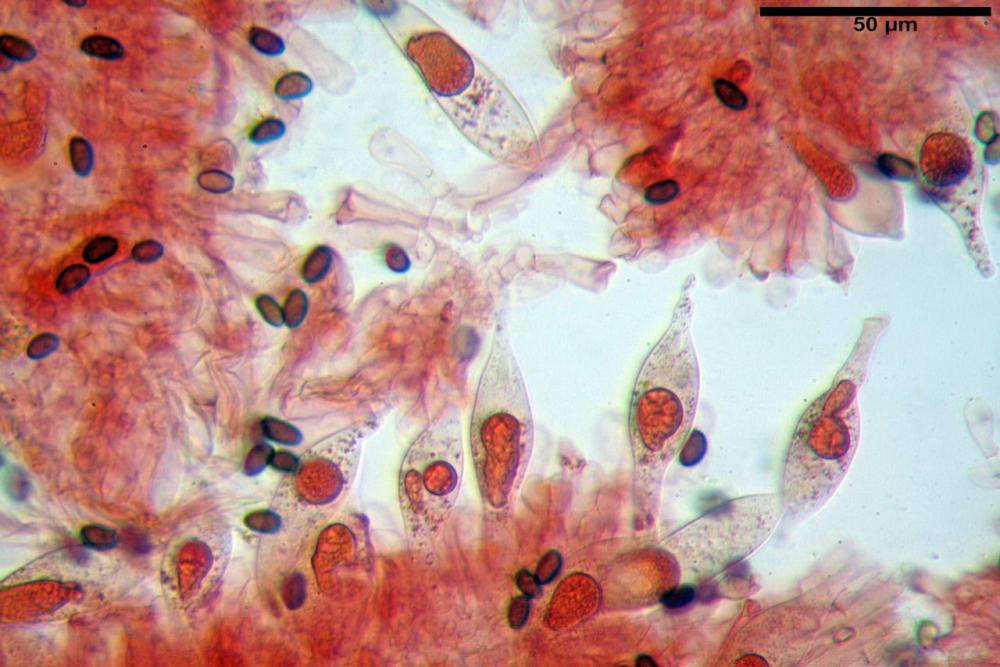
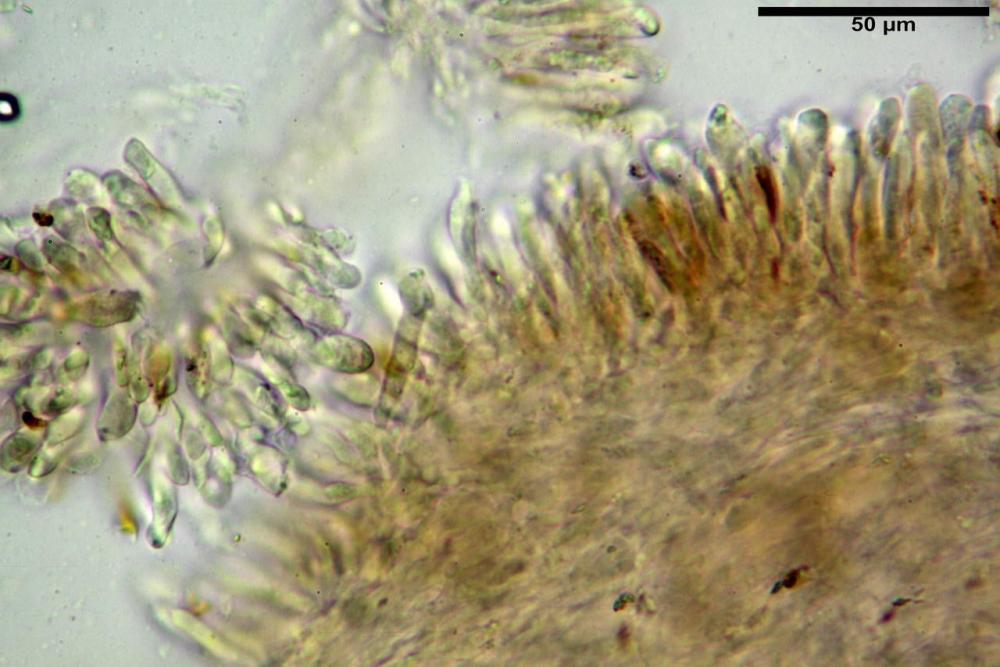
Caulocistidi (37,1)40,1-80,3(100,5) x (7,2)7,6-20,0(22,1) µm, in media 62,3 x12,9 µm, simili ai cheilocistidi ma più spesso con apice bifido (Opuntia), bitorzoluti, disposti a ciuffi o sparsi sulla superficie della caulopellis, più fitti nella zona superiore dello stipite, rarefatti verso la base.
Giunti a fibbia presenti in tutti i tessuti del basidioma.
Reazioni microchimiche negativa con l'ammoniaca (non produce cristalli nella zona imeniale).
Materiale studiato ed ambiente della raccolta: F.V.G. Trieste. Località: Trieste città Ponziana. Coordinate geografiche: 45°38'11"N 13°46'48"E. Altezza 19 m s.l.m. Habitat: su un profondo strato di residui legnosi misto a terriccio. Raccolta: 13.4.2018 (n°7474). Leg. Picciola & det. Zugna.
Osservazioni
Il ritrovamento di questa graziosa Pholiotina in ambiente fortemente antropizzato ha suscitato in noi una certa sorpresa poiché finora l’abbiamo sempre rinvenuta su residui legnosi in ambienti igrofili, come margini di stagni, boschetti golenali, etc.
Si tratta di una Pholiotina dalle caratteristiche peculiari, caratterizzata dalla presenza di deboli residui velari al margine del cappello (lente!), il gambo inizialmente bianco candido, interamente pruinoso, le colorazioni aranciate della superficie pileica negli esemplari ben idratati.
In seno al suo genere di appartenenza, sul piano microscopico Pholiotina dasypus possiede una caratteristica unica contrassegnata da pore minutamente verrucose (osservazione in olio da immersione!), con presenza di callus porico piuttosto che di un visibile poro germinativo.
Inoltre, il quadro microscopico si completa per i cheilocistidi perlopiù utriformi fino a clavati.
Si tratta di una specie alquanto variabile per quanto riguarda le dimensioni dei basidiomi, delle pore, del poro germinativo e dei cheilocistidi.
Pholiotina dasypus

Pholiotina dasypus pileipellis
Pholiotina dasypus cheilocistidi
Pholiotina dasypus caulocistidi
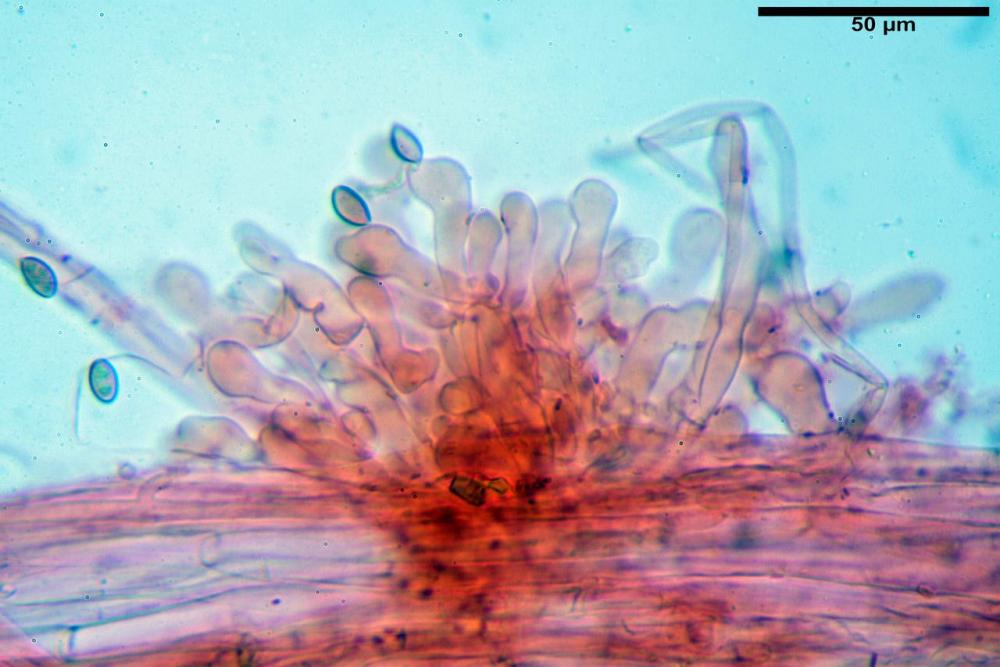
Pholiotina dasypus spore